

Глава_4
.pdfГлава 4
ПОЛОВЫЕ КЛЕТКИ. ЭМБРИОНАЛЬНОЕ РАЗВИТИЕ
Цель занятия:
Сформировать представление о строение половых клеток и об основных событиях, происходящих в половых клетках во время гаметогенеза. Усвоить механизм наследования генетического материала. Ознакомиться с основными этапами эмбриогенеза на примере развития ланцетника и лягушки. Разобрать особенности оплодотворения, дробления, гаструляции, нейруляции.
Задачи занятия:
1.Ознакомиться со стадиями овогенеза и основными процессами, происходящими в них.
2.Ознакомиться со стадиями сперматогенеза и основными процессами, происходящими в них.
3.Изучить морфологию половых клеток, связанную с выполняемыми ими функциями.
4.Рассмотреть процесс мейоза, ознакомиться с основными процессами, происходящими с хромосомами и клеткой на разных стадиях мейоза.
5.Разобрать и зарисовать основные стадии развития ланцетника по муляжам.
6.Ознакомиться с различными типами дробления, гаструляции и образования мезодермы.
7.Построить графическое изображение происхождения органов и тканей из зародышевых листков.
8.Исследовать двигательную активность сперматозоидов лягушки.
9.Изучить микропрепараты и зарисовать половые клетки животных и ранние стадии эмбриогенеза у лягушки.
Формируемые компетенции:
ОК-7, ОК-8. ОК-12, ПК-22, ПК-23
Студент должен знать:
1.Морфологию гамет, связанную с выполняемой ими функцией.
2.Название стадий оогенеза и сущность происходящих в них процессов.
3.Название стадий сперматогенеза и сущность происходящих в них процессов.
4.Отличие оогенеза и сперматогенеза.
2
5.Особенность гаметогенеза у человека.
6.Цитологическую характеристику фаз первого и второго деления мейоза.
7.Отличие мейоза и митоза.
8.Биологическое значение мейоза.
9.Аномалии числа и структуры хромосом, возникающие в ходе мейоза.
10.Классификацию яйцеклеток в зависимости от количества и распределения в них желтка.
11.Процессы, происходящие в клетках и с клетками во время эмбрионального периода: оплодотворения, периода образования зиготы, дробления, гаструляции, образования зародышевых листков, нейруляции, гисто- и органогенеза на примере ланцетника.
12.Типы дробления.
13.Виды бластул в зависимости от типа яйцеклеток.
14.Способы гаструляции.
15.Способы образования мезодермы.
16.Механизм нейруляции.
17.Провизорные органы и их функции.
18.Понятие о некоторых механизмах онтогенеза: ооплазматическая сегрегация, дифференциальный рост, размножение и гибель клеток, специализация клеток, эмбриональная индукция, дифференциальная активность генов.
19.Понятие о тератогенных факторах, вызывающих аномалии развития.
20.Иметь представление о критических периодах развития.
Студент должен уметь:
1.По описанию и по характерным признакам различать на микрофотографиях и рисунках половые клетки разных животных и клетки, находящиеся на разных стадиях гаметогенеза и мейоза.
2.Объяснить причины возникновения хромосомных аномалий
3.Идентифицировать различные стадии раннего эмбриогенеза по микропрепаратам, микрофотографиям, муляжам, рисункам и схемам
4.Зарисовывать изучаемые муляжи и микропрепараты.
Студент должен владеть:
1.Медико-функциональным понятийным аппаратом и терминологией по данной теме.
2.Навыками распознавания половых клеток разных животных.
3.Навыками диагностики структур на разных стадиях раннего эмбриогенеза на гистологических микропрепаратах.
4.Навыками устного и письменного описания стадий раннего эмбриогенеза.
Оснащение занятия:
1.Таблицы:
1.мейоз
2.гаметогенез (овогенез и сперматогенез)
3.типы дробления
4.типы гаструляции
5.развитие ланцетника
2.Микроскопы:
3
1.прямой световой монокулярный
2.прямой световой бинокулярный
3. Микропрепараты:
1.сперматозоид морской свинки
2.яйцеклетка кошки
3.мейоз яйцеклетки конской аскариды
4.эмбриогенез лягушки
4. Муляжи стадий развития ланцетника.
5. Живые сперматозоиды лягушки
Хронологическая карта занятия:
1.Организационная часть
2.Тестовый контроль базового уровня знаний
3.Объяснение практического задания
4.Самостоятельная работа
5.Проверка выполненных работ в альбомах
6.Контроль конечного уровня знаний
7.Установка задания для подготовки к следующей теме
Теоретический обзор
Мужские и женские половые клетки
Сперматозоид
Открытие сперматозоидов принадлежит голландскому натуралисту Антони ван Левенгуку. В 1679 г. Левенгук доложил о «семенных зверьках» в семенной жидкости человека Британскому Королевскому научному обществу. Термин «сперматозоид» ввёл академик Петербургской Академии Карл Эрнст фон Бэр в начале XIX века.
В сперматозоиде человека различают головку, тело и хвостик/жгутик (flagellum) (рис. 4-1). Яйцевидная, сплющенная с двух сторон головка содержит ядро с гаплоидным набором хромосом (22+Х, 22+Y) и акросому с ферментами для растворения прозрачной оболочки яйцеклетки (гиалуронидаза, протеазы, гликозидазы, липазы, нейраминидаза, фосфатазы). Тело сперматозоида имеет утолщение за счёт спирально закрученной нитевидной митохондрии. Между телом и головкой расположены проксимальная (центр организации аксонемы) и дистальная (имеющая кольцевую форму) центриоли. В осевой части тела и хвостика, начиная от проксимальной центриоли, находится сократительная органелла — аксонема, образованная микротрубочками по принципу 9+2 (девять пар микротрубочек по периферии и одна пара в центре). Снаружи от аксонемы расположено 9 фибрилл, состоящих из электроноплотного материала. Фибриллы начинаются от шейки, проходят через тело и заканчиваются на уровне середины хвостика. Фибриллы в пределах хвостика окружены фибриллярным футляром из электроноплотного материала, не доходящего до концевого отдела хвостика.
4
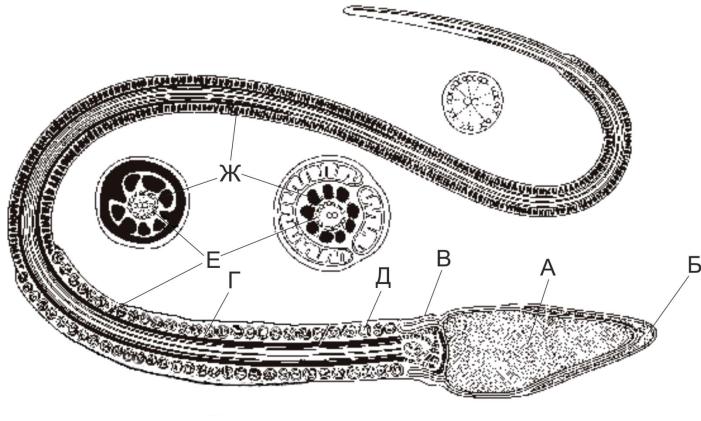
Рис. 4-1. Сперматозоид человека. Головка содержит ядро (А) и акросому (Б). В шейке (В) присутствуют дистальная и проксимальная центриоли. Промежуточный отдел (Г) содержит спирально закрученную нитевидную митохондрию (Д). В осевой части промежуточного отдела и почти вдоль всего хвоста расположена аксонема (Е). В хвосте снаружи от неё находятся наружные плотные фибриллы и фибриллярный футляр (Ж) [из: Fawcett D.W., 1975].
В эякуляте здорового мужчины содержится примерно от 150 до 600 млн сперматозоидов. В 23% случаев мужского бесплодия обнаруживается патозооспермия (отсутствие или недостаточное количество сперматозоидов, морфологические дефекты, снижение подвижности сперматозоидов).
•Спермограмма включает характеристики сперматозоидов (количество, морфология, жизнеспособность, подвижность, агглютинация) и семенной жидкости (объём, рН, вязкость, лейкоциты, микрофлора).
Яйцеклетка
Женская половая клетка имеет шарообразную форму диаметром 100-150 µм, не способна к активному движению. Центрально расположенное округлое ядро с выраженным ядрышком содержит гаплоидный набор хромосом (22+Х). В цитоплазме яйцеклетки, кроме типичных органелл, содержится небольшое количество желточных включений (lecythos) с питательными веществами (белки, жиры, углеводы, минеральные соли), используемыми на начальных этапах развития зародыша, а под плазмалеммой локализуются кортикальные гранулы, содержащие ферменты (протеазы).
5
Сегрегационные гены
Яйцеклетка гетерогенная, химически преформированная высокоспециализированная клетка организма. Томас Морган предложил процесс созревания яйцеклетки считать точкой отсчёта онтогенеза, поскольку именно в ходе созревания яйцеклетки закладывается план будущего строения организма. Ядро развивающегося овоцита работает с опережением (на будущее). В цитоплазме накапливаются самые разнообразные мРНК, специфически распределённые в цитоплазме. В результате ооплазматической сегрегации возникает полярный градиент распределения биологически активных веществ. В яйцеклетке можно выделить три системы генов материнского эффекта, контролирующих формирование трёх градиентов: анимальновегетативного, дорсо-вентрального и формирования головных и хвостовых структур. Сегрегационные гены материнского эффекта контролируют активность gap-генов, pair-rule генов и генов сегментарной полярности, последовательно осуществляющих сегментацию зародыша.
Цитоплазматический контроль активности хромосом. Клонирование
Функциональное состояние ядра зависит от вида цитоплазмы, в которой находится ядро. Ядро выдаёт наследственную информацию в соответствии с цитоплазматическим окружением. Переход ядра в новое функциональное состояние характеризуется накоплением в ядре цитоплазматических белков, в результате чего ядра в разных клеточных типах проявляются дифференциальной экспрессией генов. Обратимость изменений в ядрах дифференцированных соматических клеток была установлена в 60-е годы ХХ в. Джон Гердон (Оксфорд, Великобритания) трансплантировал ядро клетки кишечника взрослых лягушек в облучённое яйцо лягушки. Около 1% таких яиц развивался во взрослых лягушек. Клонирование стало общеизвестным в 1997 г., когда Ян Вильмут (Эдинбург, Шотландия) клонировал овцу Долли.
Гаметогенез
Мужские и женские половые клетки (гаметы) образуются в ходе гаметогенеза.
Развитие мужских половых клеток — сперматозоидов (сперматогенез) совершается в мужских половых железах — яичках, развитие женских половых клеток — яйцеклеток (овогенез) происходит в яичниках — женских половых железах.
Сперматогенез (образование сперматозоидов) подразделяют на четыре стадии: (1) размножения, (2) роста, (3) созревания (мейоз) и (4) формирования (спермиогенез). Первичные половые клетки мигрируют в зачатки яичек, делятся и дифференцируются в сперматогонии. До периода полового созревания сперматогонии остаются в состоянии покоя. Стадия размножения начинается с наступлением половой зрелости. После ряда митотических делений сперматогонии дифференцируются в сперматоциты первого порядка, вступающие в стадию роста. Сперматоциты увеличиваются в размерах в 4 и более раз. Стадия созревания (мейоз) следует сразу за стадией роста. В результате первого деления мейоза из одного сперматоцита первого порядка образуется два сперматоцита второго порядка, а после второго мейотического деления — четыре сперматиды, имеющие по 22 аутосомы и одной X- или Y-хромосоме. Сперматоциты второго порядка в два раза, а сперматиды в четыре раза меньше по объёму спематоцитов первого порядка. Спермиогенез (стадия формирования) — постмейотическая стадия морфологических изменений сперматид с образованием сперматозоидов (рис. 4-2). Таким образом, в ходе сперматогенеза из одной сперматогонии образуется четыре полноценных сперматозоида.
6
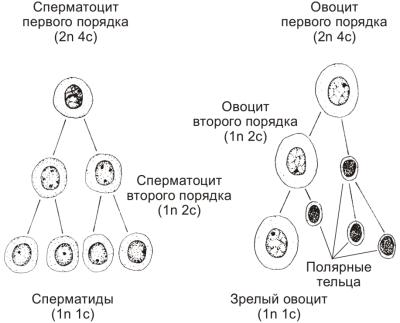
Овогенез (образование яйцеклетки) проходит через три стадии: (1) размножения, (2) роста и (3) созревания (мейоз). Первичные половые клетки мигрируют в зачатки яичников и дифференцируются в овогонии, которые сразу вступают в стадию размножения. Завершив серию митотических делений, овогонии вступают в стадию роста. В этот период в цитоплазме накапливаются желточные включения. Вслед за стадией роста начинается стадия созревания (мейоз). Первое деление мейоза остается незавершённым: образующиеся овоциты первого порядка в профазе первого деления мейоза вступают в длительный период покоя, продолжающийся до наступления половой зрелости. С наступлением половой зрелости и установлением овариальноменструального цикла при овуляции (выхода яйцеклетки из фолликула) завершается первое деление мейоза и начинается второе деление, останавливающееся в метафазе. При этом образуеся крупный овоцит второго порядка и мелкая абортивная клетка — первое полярное (направительное, или редукционное) тельце. Сигнал для завершения второго мейотического деления — оплодотворение; овоцит второго порядка делится с образованием зрелой яйцеклетки и второго полярного тельца. Первое полярное тельце также подвергается второму делению мейоза. Зрелая яйцеклетка имеет 22 аутосомы и одну X-хромосому (рис. 4-2). Таким образом, в ходе овогенеза из одной овогонии образуется одна полноценная яйцеклетка, под прозрачной оболочкой которой локализуются три полярных тельца.
Рис. 4-2. Созревание мужских и женских половых клеток. Перед первым мейотическим делением генетический материал удваивается с образованием конъюгированных хромосом (2n4c). После первого деления мейоза в дочерних клетках уменьшаются количество хромосом и содержание ДНК; остаётся по 23 конъюгированных (удвоенных) хромосомы с диплоидным содержанием ДНК (1n2c). После второго деления мейоза дочерние клетки получают по 23 хромосомы с гаплоидным содержанием ДНК (1n1c) — 22 аутосомы и одну половую хромосому. n — число хромосом, c — количество ДНК [из: Kaplan S., 1987].
7

Мейоз. В ходе мейоза образуются гаплоидные гаметы (рис. 4-3). При этом происходят следующие события:
•генетическая рекомбинация путём кроссинговера между гомологичными хромосомами (отцовскими и материнскими),
•уменьшение числа хромосом,
•снижение содержания ДНК,
•уменьшение плоидности клеточных потомков,
•значительный синтез РНК.
Рис. 4-3. Мейоз обеспечивает переход половых клеток из диплоидного состояния в гаплоидное. Клетка, вступающая в мейоз, проходит два деления: первое деление — редукционное, второе
— эквационное [из: Gilbert S.F., 1985].
8
Первое деление мейоза
Первое деление мейоза (профаза I, метафаза I, анафаза I и телофаза I) — редукционное.
Профаза I. Профаза I последовательно проходит несколько стадий: лептотена, зиготена, пахитена, диплотена, диакинез.
•Лептотена. Хроматин конденсируется, каждая хромосома состоит из двух хроматид, соединённых центромерой.
•Зиготена. Гомологичные парные хромосомы сближаются и вступают в физический контакт (синапсис) в виде синаптонемального комплекса, обеспечивающего конъюгацию хромосом. Контакт позволяет хромосомам обмениваться генетическим материалом (кроссинговер). На этой стадии две лежащие рядом пары хромосом образуют бивалент.
•Пахитена. Хромосомы утолщаются вследствие спирализации. Отдельные участки конъюгировавших хромосом перекрещиваются друг с другом и образуют хиазмы. Здесь происходит кроссинговер — обмен участками между отцовскими и материнскими гомологичными хромосомами, что определяет генетические различия между индивидуумами.
•Диплотена. Происходит разделение конъюгировавших хромосом в каждой паре в результате продольного расщепления синаптонемального комплекса. Хромосомы расщепляются по всей длине комплекса, за исключением хиазм. В составе бивалента чётко различимы 4 хроматиды. Такой бивалент называют тетрадой. В хроматидах появляются участки раскручивания, где синтезируется РНК.
•Диакинез. Продолжаются процессы укорочения хромосом и расщепления хромосомных пар. Хиазмы перемещаются к концам хромосом (терминализация). Разрушается ядерная мембрана, исчезает ядрышко. Появляется митотическое веретено.
Метафаза I. В метафазе I тетрады образуют метафазную пластинку. В целом отцовские и материнские хромосомы распределяются случайным образом по ту или другую сторону экватора митотического веретена. Подобный характер распределения хромосом лежит в основе второго закона Менделя, что (наряду с кроссинговером) обеспечивает генетические различия между индивидуумами.
Анафаза I. Анафаза I отличается от анафазы митоза тем, что при митозе к полюсам расходятся сестринские хроматиды. В эту фазу мейоза к полюсам отходят целостные хромосомы.
Телофаза I. Телофаза I не отличается от телофазы митоза. Формируются ядра, имеющие 23 конъюгированных (удвоенных) хромосомы, происходит цитокинез, образуются дочерние клетки.
Второе деление мейоза
Второе деление мейоза — эквационное — протекает так же, как митоз (профаза II, метафаза II, анафаза II и телофаза II), но значительно быстрее. Дочерние клетки получают гаплоидный набор хромосом (22 аутосомы и одну половую хромосому).
Аномалии хромосом
Хромосомные нарушения возникают в ходе мейоза de novo, поэтому родители могут быть здоровы по этому признаку.
Аномалии числа хромосом возникают вследствие нерасхождения хромосом в мейозе (рис. 4-4).
•Половые хромосомы. Синдром Тернера (45Х0) встречается с частотой 1:2500, синдром Кляйнфелтера (47ХХY) 1:1000, 47ХХХ 1:1000, 47ХYY 1:1000.
9
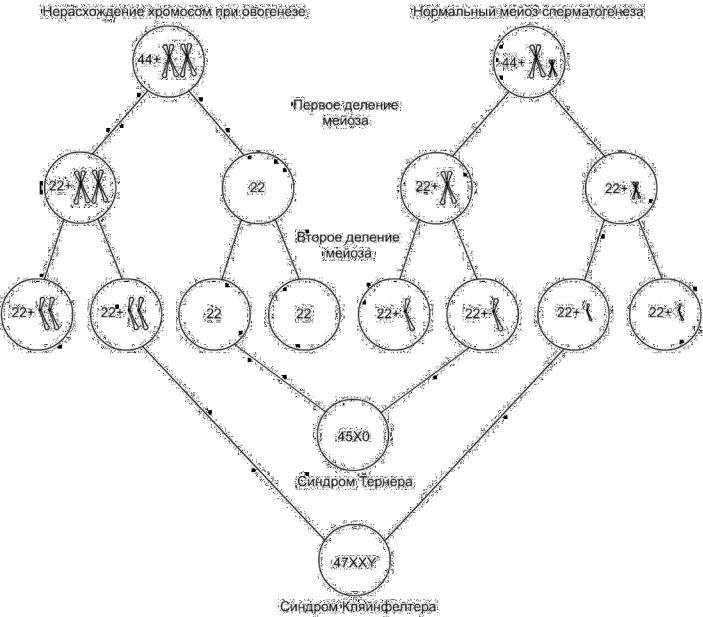
•Аутосомы. Синдром Дауна (трисомия 21) встречается с частотой 1:700, синдром Эдвардса (трисомия 13) 1:4000), синдром Патау (трисомия 18) 1:8000.
Рис. 4-4. Нерасхождение хромосом в мейозе.
Аномалии структуры хромосом. В ходе кроссинговера в хромосомах, обменивающихся гомологичными участками, возникает перераспределение или утеря части генного материала: инверсии, делеции, дупликации, транслокации.
•Транслокация гена sry, кодирующего TDF (Testis Determing Factor), из Y-хромосомы в X- хромосому в ходе кроссинговера приводит к изменению пола и дисгенезии гонад. В случае
10
оплодотворения яйцеклетки сперматозоидом с Y-хромосомой, утратившей ген sry, генетические мужчины (46XY) имеют женский фенотип и, наоборот, при оплодотворении яйцеклетки X-хромосомой, получившей ген sry, генетические женщины (46XX) имеют мужской фенотип.
Эмбриональное развитие
Основные понятия. Эмбриональным развитием, или эмбриогенезом, называют начальный период онтогенеза; это процесс развития многоклеточного организма из оплодотворённой яйцеклетки, т.е. от зиготы до момента рождения на свет нового организма. В этот период последовательно протекают стадии дробления, гаструляции, гисто- и органогенеза, формирования пространственной организации тела (морфогенез). Одновременно координированным образом происходит пролиферация (размножение), рост, миграция (перемещение), детерминация (определение возможного пути дифференцировки), дифференцировка (специализация к выполнению конкретной функции) и апоптоз (гибель) клеток, обеспечивающих реализацию эндогенной программы развития многоклеточного организма из одной единственной клетки (зиготы).
Зигота. Слияние мужской и женской гамет (оплодотворение) приводит к образованию одноклеточного зародыша — зиготы. При оплодотворении взаимодействуют мужская и женская гаплоидные гаметы; при этом сливаются их ядра (пронуклеусы), объединяются хромосомы и возникает диплоидная клетка нового организма — зигота. Начало оплодотворения — момент слияния мембран сперматозоида и яйцевой клетки, окончание оплодотворения — момент объединения материала мужского и женского пронуклеусов.
Митохондриальный геном
Митохондриальная ДНК наследуется по материнскому типу. В яйцеклетке количество митохондрий составляет несколько тысяч. В сперматозоиде человека присутствует только одна спирально закрученная митохондрия, которая подвергается деградации и перевариванию в зиготе.
Дробление — непрерывные митотические деления диплоидных клеток (зиготы и бластомеров) без последующего роста дочерних клеток, отчего объём зародыша не меняется. Характер дробления и образующейся в итоге тип бластулы определяется количеством и распределением в цитоплазме яйцеклетки желточных включений:
-изолецитальная яйцеклетка обусловливает полное равномерное дробление. Бластомеры делятся полностью на равные по размерам дочерние клетки (голобластический тип дробления). При синхронном типе дробления соблюдается геометрическая прогрессия, последовательно образуется 2, 4, 8, 16. 32, 64, 128 бластомеров. При асинхронном типе дробления встречается, например, 3- клеточная стадия;
-оплодотворённая телолецитальная яйцеклетка также делится целиком, однако на анимальном полюсе, где клетки содержат мало желтка, дробление происходит быстрее и образуются многочисленные мелкие клетки. Напротив, на вегетативном полюсе локализуются крупные
11