

Глава_4
.pdfнагруженные желтком клетки. Здесь дробление происходит медленнее, поэтому образуется меньшее количество более крупных клеток (полное неравномерное дробление);
-в оплодотворённых резко телолецитальных и центролецитальных яйцеклетках дробится цитоплазма, свободная от желтка (неполное, частичное дробление, или меробластический тип дробления);
-дискоидальное неполное дробление характеризуется дроблением цитоплазмы, содержащей ядро, в области анимального полюса. При поверхностном неполном дроблении дробится слой цитоплазмы, окружающий снаружи центральное скопление желточных гранул.
По типу симметрии дробления оплодотворённой яйцеклетки различают:
-радиальное (ось яйцеклетки является осью радиальной симметрии делящихся бластомеров);
-спиральное (бластомеры, возникающие в результате последующих делений, располагаются друг относительно друга по спирали);
-билатеральное (бластомеры располагаются таким образом, что у зародыша имеется только одна плоскость симметрии);
-анархичное (у одного вида встречаются разные варианты расположения бластомеров, бластомеры образуют цепочки или бесформенную клеточную массу).
•Морула. По мере увеличения числа бластомеров в результате компактизации происходит уменьшение размеров межклеточных пространств между бластомерами, они сближаются и формируется морула. Морула — эмбриологический термин, введённый Геккелем (Heckel) для обозначения заключительной стадии дробления, когда зародыш представляет собой компактное скопление бластомеров перед формированием бластулы.
•Бластула. Эта стадия развития завершает период дробления. Строение зародыша на стадии бластулы зависит от строения яйцеклетки и типа дробления (рис. 4-5).
o В результате полного равномерного дробления формируется целобластула, представляющая собой полый шар, стенка которого образована одним слоем клеток (бластодерма). Полость (бластоцель), заполненная жидкостью, занимает весь внутренний объём бластулы.
o Амфибластула образуется из телолецитальной яйцеклетки в результате полного неравномерного дробления. Бластодерма состоит из неодинаковых по объёму бластомеров анимального (мелких клеток) и вегетативного (крупных клеток) полюсов. Бластоцель смещён к анимальному полюсу.
o Дискобластула представлена дисковидным скоплением клеток, отделённых щелевидной полостью (бластоцель) от нераздробившегося желтка. Образуется из резко телолецитальной яйцеклетки в результате неполного дискоидального дробления.
o Перибластула образуется из центролецитальной яйцеклетки в результате поверхностного неполного дробления. Стенка перибластулы состоит из одного слоя клеток, а полость заполнена нераздробившимся желтком.
o Стерробластула (от греч. sterros — твёрдый, плотный), образуется обычно в результате спирального дробления. Она имеет вид плотного шара, не содержащего бластоцель.
12

Рис. 4-5. Типы бластул. А — целобластула; Б — амфибластула; В — дискобластула; Г — перибластула; Д
— стерробластула [из: Кнорре А.Г., 1959, по Кацнельсону З.С.].
Гаструляция — следующая за дроблением стадия эмбриогенеза, характеризующаяся появлением у клеток способности к росту и перемещению. За счёт активной пролиферации, роста, направленной миграции и дифференцировки клеток бластодермы из бластулы образуются трёхслойный зародыш (гаструла) с первичными зародышевыми листками: эктодермой, мезодермойа и энтодермой. В зависимости от типа бластулы гаструляция происходит различными способами, или может быть результатом комбинации двух и более разных способов (рис. 4-6). Обычно сначала образуется двухслойная гаструла, стенка которой представлена наружным слоем клеток — эктодермой и внутренним слоем клеток — энтодермой. Новая формирующаяся полость (гастроцель) сообщается с внешней средой через отверстие — бластопор (первичный рот). Несколько позже образуется третий зародышевый листок — мезодерма. Клетки зародышевой мезодермы формируют дорсальную мезодерму, из которой возникают хорда — продольный плотный тяж клеток и сомиты — симметричные парные структуры по бокам от хорды и нервной трубки, состоящие из клеток дерматома, миотома и склеротома. Из мезодермы образуется ещё два крупных зачатка: промежуточная мезодерма (нефротом) и латеральная мезодерма (спланхнотом), расщепляющаяся на париетальный и висцеральный листки, ограничивающие целомическую полость.
13

Рис. 4-6. Способы гаструляции. А — инвагинация происходит путём впячивания вегетативного полушария (будущая энтодерма) бластулы в бластоцель под её анимальный полюс (будущая эктодерма); Б
— эпиболия происходит путём обрастания вегетативного полюса бластулы быстро делящимися клетками анимального полюса. В результате клетки анимального полюса формируют наружный листок — эктодерму, а клетки вегетативного полюса — энтодерму; В — иммиграция — выселение клеток бластодермы в бластоцель с образованием внутреннего слоя клеток (энтодермы). Клетки, оставшиеся на месте, формируют эктодерму. При униполярной (однополюсной) иммиграции выселение клеток происходит на вегетативном полюсе, при мультиполярной (многополюсной) — в нескольких местах бластулы; Г — деламинация характеризуется расслоением бластодермы на два слоя клеток; наружный слой преобразуются в эктодерму, а внутренний слой формирует энтодерму [из: Кнорре А.Г., 1959, по Кацнельсону З.С.].
14
Морфогенез — формирование пространственной организации частей организма. Морфогенез осуществляется при реализации различных морфогенетических процессов. Под контролем сигнальных молекул морфогенов (из семейства гомейозисных генов) создаётся морфогенетическое поле. Клетки, занявшие определённое положение в системе развивающегося зародыша (морфогенетическое поле), получают позиционную информацию и приступают к выполнению программы детерминации и дифференцировки (морфогенетическая реакция, в результате которой происходит качественная спецификация сегментов: формообразование органов, систем, частей тела). Активность сегрегационных генов яйцеклетки обеспечивает нормальное функционирование системы гомейозисных генов, контролирующих качественную спецификацию сегментов. Под гомейозисом, согласно автору этого термина генетика Уильяма Бэтсона (1984 г.), следует понимать превращение одной части тела в другую. Мутация гена, контролирующего развитие определённого сегмента, вызывает его превращение в «двойника» впереди лежащего сегмента.
•Гомейозисные гены — семейство родственных генов, содержащих гомеобокс и определяющих форму тела. У млекопитающих это семейство представлено 38 генами,
сгруппированными в 4 комплекса — Hox (Homeobox) A, Hox B, Hox C и Hox D, или , по номенклатуре Human Gene Mapping Workshops (рабочая группа картирования генов человека), соответственно Hox 1, Hox 2, Hox 3 и Hox 4. Гены экспрессируются в эмбриогенезе и определяют организацию общего плана тела.
•Гомеобокс — эволюционно консервативная последовательность, состоящая примерно из 180 пар нуклеотидов. Соответствующая последовательность из 60 аминокислотных остатков, кодируемая гомеобоксом, получила название гомеодомен. Гены, содержащие гомеобокс, кодируют ядерные белки, регулирующие экспрессию генов, а гомеобокс кодирует часть ДНК-связывающего белка.
Нейруляция— процесс закладки нервной системы. В результате индуцирующего влияния друг на друга зародышевых листков начинается образование новых структур (морфогенез). Первичная эмбриональная индукция (влияние хордомезодермы на дорсальную эктодерму) инициирует
процесс первичного органогенеза формирования нервной трубки, дающей начало нервной системе на месте презумптивной нейроэктодермы (рис. 4-7).
•Молекулярные сигналы нейруляции. На третьей неделе развития во всей нервной пластинке отмечена экспрессия гомеобокс-содержащих генов PAX3 и PAX7. На этапе формирования нервных желобков секретируемый сначала хордой, а затем вентральной частью нервной трубки sonic hedgehog сдерживает экспрессию PAX3 и PAX7 в вентральной части нервной трубки. Экспрессию PAX3 и PAX7 в дорсальной части нервной трубки поддерживают морфогенетические белки кости (BMP4 и BMP7) из ненейрогенной эктодермы.
•Морфогены семейства hedgehog. При мутации гена hh эмбрион дрозофилы покрывается шипиками, напоминая ёжика, за что ген и получил своё название (hedgehog от англ. ёжик). У млекопитающих были открыты три гомологичных гена (desert hedgehog, indian hedgehog и sonic hedgehog). Название гена sonic hedgehog заимствовано из популярной Sega видеоигры от персонажа Hedgehog the Sonic. Названия генов desert hedgehog и indian hedgehog соответствуют двум видам ёжиков.
15

Рис. 4-7. Нейруляция. В ходе нейруляции последовательно происходят следующие стадии: формирование нервной пластинки → приподнимание краёв нервной пластинки и образование нервного желобка → появление нервных валиков → формирование нервного гребня и начало выселения из него клеток → смыкание нервных валиков с образованием нервной трубки → срастание эктодермы над нервной трубкой
[из: Moore K.L., Persaud T.V.N., 1998].
Гистогенез и органогенез. Согласно клональной теории развития, любая ткань и орган берут начало из небольшой группы клонов, каждый из которых образуется из своей стволовой клетки. Закладка и развитие органов запускаются ткане- и органоспецифическими морфогенами, активно поддерживающими процессы роста и становления функций органов в заданном морфогенетическом поле. Зародышевые листки (эктодерма, энтодерма и мезодерма) дают начало всем четырём типам тканей (эпителиальной, соединительной, мышечной и нервной), из которых, в свою очередь, формируются органы и системы органов (рис. 4-8). Одной из первых закладывается и начинает функционировать сердечно-сосудистая система.
16

Рис. 4-8. Производные первичных зародышевых листков.
Практическая часть
Микропрепараты. Половые клетки. Ранний эмбриогенез лягушки
1. Сперматозоид
Сперматозоиды разных видов животных имеют похожее строение с некоторыми отличиями (рис. 4-9). Например, форма головки может быть шарообразной, как у сперматозоида морской свинки, серповидной, как у сперматозоида петуха. По количеству жгутиков встречаются сперматозоиды, имеющие несколько жгутиков (некоторые ракообразные) или не имеющие жгутика. Сперматозоиды круглых червей имеют амебоидную форму и передвигаются с помощью псевдоподий.
17

Рис. 4-9. Сперматозоид морской свинки. В округлой головке клетки чётко различимы ядро и акросома. От шейки начинается утолщённая часть хвостика — тело сперматозоида, продолжающееся в хвостовую нить.
2. Яйцеклетка
Размер и строение яйцеклетки зависят от количества и распределения желтка в цитоплазме. Алецитальная яйцеклетка не содержит желтка, встречается у плоских червей. Яйцеклетка млекопитающих содержит мало желтка (олиголецитальная) (рис. 4-10). Мезолецитальная яйцеклетка содержат среднее количество желтка (осетровые рыбы, амфибии), а полилецитальная — большое количество (членистоногие, рыбы, рептилии, птицы). В олиголецитальных яйцеклетках желток распределён равномерно (гомо-, изолецитальный тип). В полилецитальных и мезолецитальных яйцеклетках желток может быть смещён к одному из полюсов (вегетативному), тогда как противоположный полюс яйцеклетки (анимальный) свободен от желтка — такие яйцеклетки относятся телолецитальному типу. Центролецитальный тип яйцеклетки характерен для полилецитальных клеток, в которых желток расположен в центре яйцеклетки.
На микропрепарате (яичник кошки) под малым увеличением микроскопа найти фолликул, содержащий яйцеклетку (рис. 4-10). Под большим увеличением рассмотреть фолликул, различить и обозначить на рисунке структуры яйцеклетки: лучистый венец (оболочка из фолликулярных клеток), прозрачную оболочку, плазмолемму, желточные гранулы в цитоплазме, ядро с ядрышком.
18

Рис. 4-10. Яйцеклетка млекопитающего. Яичник содержит многочисленные фолликулы (пузырьки), содержащие яйцеклетки (овоциты). В зрелом фолликуле овоцит первого порядка находится внутри возвышения — яйценосного бугорка (cumulus oophorus). Кнаружи от плазматической мембраны овоцита видна толстая прозрачная оболочка (zona pellucida), к которой примыкает несколько слоёв фолликулярных клеток, образующих лучистый венец (corona radiata). Крупное ядро яйцеклетки имеет округлую форму и занимает центральное положение. В ядре хорошо заметны ядрышки. Цитоплазма овоцита содержит распределённые равномерно немногочисленные желточные включения и примембранные кортикальные гранулы [из: Gilbert S.F., 1985].
3. Мейоз яйцеклетки конской аскариды
На препарате представлен поперечный срез через зону роста половой трубки самки конской аскариды (круглый червь). Видны овоциты, покрытые трёхслойной оболочкой (внутренний слой
— липидный, наружный — белочный и средний — хитиновый) (рис. 4-3, рис. 4-11). В стадии пролиферации и роста диплоидный набор овоцита состоит из двух пар гомологичных хромосом.
Овоцит 1 порядка в профазе первого деления мейоза. На этой стадии мейоза каждая хромосома уже удвоилась и состоит из двух хроматид, а гомологичные хромосомы конъюгируют с образованием тетрад. Ядерная оболочка овоцита распалась, на месте ядра видны две тетрады. Сперматозоид уже проник в овоцит, его пронуклеус достигает центра клетки, где остается в покоящемся состоянии до окончания созревания яйцеклетки. Пронуклеус сперматозоида содержит две хромосомы и имеет вид тельца с расплывчатыми контурами.
Овоцит 2 порядка. Произошло первое (редукционное) деление мейоза. В овоците 2 порядка остались две негомологичные хромосомы, каждая из которых состоит из двух хроматид. Две другие хромосомы, также состоящие из двух хроматид, вместе с небольшим участком цитоплазмы овоцита образуют первое направительное тельце, которое отделяется от овоцита и остаётся под его оболочкой. В центре клетки виден пронуклеус сперматозоида.
Зрелая яйцеклетка. Произошло второе (эквационное) деление мейоза. В о воците остаются две хроматиды, представляющие гаплоидный набор хромосом, и пронуклеус сперматозоида с гаплоидным набором хромосом. Две другие хроматиды образуют второе направительное тельце, которое также остаётся под оболочкой яйцеклетки.
19
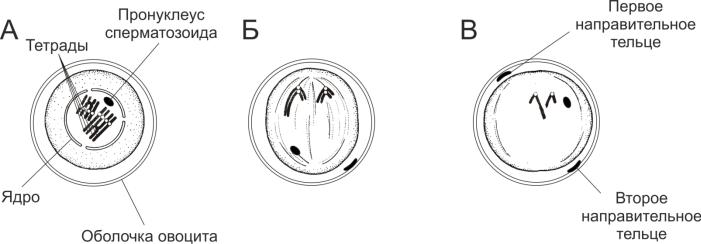
Рис. 4-11. Созревание яйцеклетки конской аскариды. А — овоцит первого порядка; Б — овоцит второго порядка; В — зрелая яйцеклетка.
4. Эмбриогенез лягушки
Микропрепарат изучить под большим увеличением. Найти и зарисовать стадии формирования бластулы (рис. 4-12).
Стадия двух бластомеров. Первая борозда дробления (меридиональная) постепенно распространяется от анимального до вегетативного полюса. Зигота делится полностью (полное дробление), в результате образуется две одинаковые по размеру дочерние клетки (равномерное дробление). При этом не происходит увеличения бластомеров и сохраняется размер зиготы.
Стадия четырёх бластомеров. Вторая борозда дробления проходит меридионально перпендикулярно первой. В результате второго деления образуется четыре одинаковых по размеру бластомера (дробление полное равномерное). По объёму бластомеры в четыре раза меньше зиготы.
Морула. Третья борозда дробления проходит по широте и смещена к анимальному полюсу, что обусловлено загруженностью желтком вегетативного полюса. На 8-клеточной стадии появляется различие в размерах бластомеров. В результате полного неравномерного дробления образуются четыре анимальных бластомера, значительно меньших по объёму четырёх вегетативных клеток. Следующие две меридиональные борозды возникают одновременно, при этом образуются 16 бластомеров. Следующие две одновременные широтные борозды приводят к образованию 32клеточной стадии. Далее чередуются четыре одновременных меридиональных и четыре одновременных широтных борозд дробления, в результате чего появляется 128-клеточный зародыш. Однако, после 64-клеточной стадии геометрическая прогрессия нарушается. Клетки анимального полюса, бедные желтком, делятся быстрее клеток вегетативного полюса, нагруженных желточными включениями.
Бластула. По мере увеличения количества бластомеров клетки расходятся и между ними формируется полость, которая заполняется жидкостью. У амфибий в результате полного неравномерного дробления образуется амфибластула. Бластоцель амфибластулы располагается асимметрично (смещён к анимальному полюсу). Бластодерма (стенка бластулы) имеет разную толщину за счёт различия в количестве слоёв и размеров бластомеров. В анимальной части, в области крыши, мелкие клетки образуют 1-3 слоя клеток. В вегетативной части, в области дна, толстая стенка образована крупными, загруженными желточными гранулами, клетками. В
20

экваториальной (краевой) зоне, между крышей и дном амфибластулы, бластодерма представлена 3-5 слоями промежуточных по размеру клетками.
Рис. 4-12. Дробление и образование бластулы лягушки [из: Кнорре А.Г., 1959].
Лабораторная работа. Двигательная активность сперматозоидов. Муляжи начальных этапов эмбриогенеза ланцетника
1. Живые сперматозоиды лягушки
Фрагмент семенника лягушки погрузить в физиологический раствор, перемешать стеклянной палочкой, слегка надавливая на семенник, в результате чего выделяется молочно-белая жидкость. Каплю полученной жидкости нанести на предметное стекло и покрыть покровным стеклом. Под малым увеличением микроскопа найти хаотично двигающиеся сперматозоиды (рис. 4-13). Обратить внимание на хлыстообразное биение хвостика сперматозоида при его движении и палочковидное строение его головки.
Рис. 4-13. Сперматозоид лягушки. Клетка имеет головку палочковидной формы и хлыстообразно изгибающийся при движении хвостик.
21