

5 Обмен аммиака. Биосинтез мочевины. Некоторые клинические аспекты
Аммиак – это один изконечных продуктов обмена азотсодержащих веществ. Это составляющая фракции остаточного азота сыворотки крови (наряду с мочевиной, мочевой кислотой, креатинином, индиканом). В крови концентрация аммиака невелика - 25-40 мкмоль/л. При более высоких концентрациях он оказывает токсическое действие на организм.
Аммиак токсичен, в первую очередь для ЦНС. Токсичность аммиака связана с его способностью нарушать функционирование ЦЛК, т.к. NH3выводит из ЦЛК–кетоглутарат:
–КГ + NH3+ НАДН.Н+Глу + НАД++ Н2О.
В итогевосстановительного аминирования –кето–глутарата происходит снижение активности ЦЛК в клетках ЦНС, что, в свою очередь, угнетает активность аэробного окисления глюкозы. В результате нарушается энергопро–дукция и развивается гипоэнергетическое состояние, т.к. глюкоза – это основной источник энергии для головного мозга.
NH3 образуется в ходе следующих процессов:
1) окислительного дезаминирования аминокислот – это основной путь продукции NH3 ;
дезаминирования биогенных аминов;
дезаминирования пуриновых оснований (аденин, гуанин);
катаболизм пиримидиновых нуклеотидов.
В головном мозге основной источник образования NH3 – дезаминирование АМФ до инозинмонофосфата (ИМФ):
АМФ + Н2О ИМФ + NH3.
Фермент, который катализирует эту реакцию, - аденозиндезаминаза.
Аммиак транспортируется кровью к печени и почкам для обезвреживания в составе аминокислот, среди которых основными являются глутамин, аспарагин, аланин.
Обезвреживание NH3происходит практически сразу после его образования, т.к. в тканях он сразу же включается в состав аминокислот, главным образом глутамина. Однако для дальнейшей детоксикации и выведения амиака существуют биохимические процессы в печени и почках, которые и являются основными путями обезвреживанияNH3.
Выделяют следующие механизмы обезвреживания NH3:
1) восстановительное аминирование–кетоглутарата;
2) образование амидов аминокислот – аспарагина и глутамина;
3) образование аммонийных солей в почках;
4) синтез мочевины.
В тканях аммиак подлежит немедленной нейтрализа–ции. Это достигается путем сочетания процессов (1) и (2).
Восстановительное аминирование –кетоглутарата:
NH3 + –КГ + НАДН.Н+ Глу + НАД + Н2О.
Фермент - глутаматдегидрогеназа
Для этого процесса необходимы значительные концентрации –КГ. Для того чтобы не было перерасхода–КГ и работа ЦЛК не была нарушена,–КГ пополняется за счет превращения ПВКОА–КГ .
2)Образование амидов– это важный вспомогательный механизм обезвреживанияNH3в тканях путем его связывания с Глу или Асп.
Асп + АТФ + NH3 Асн + АМФ + ФФнн
Фермент - аспарагинсинтаза
Глу + АТФ + NH3 Глн + АМФ + ФФнн
Фермент - глутаминсинтаза
Этот процесс наиболее активен в ЦНС, мышцах, почках, печени (для поддержания внутренней концентрации NH3). Главным образомглн является транспортной формой нетоксичногоNH3 из мозга, мышц и др. тканей. Глутамин легко проникает через мембрану, т.к. при физиологических значениях рН он не имеет заряда. При физической нагрузке аланин активно транспортируетNH3от мышц к печени. Кроме того, большое количество аланина содержит кровь, оттекающую из кишечника. Этот аланин также направляется в печень для глюконеогенеза.
3) Глн и асн с током крови попадают в почки, где подвергаются гидролизу с помощью специальных ферментов – глутаминазы и аспарагиназы, которые есть и в печени:
Асн + Н2ОАсп +NH3.
Глн + Н2ОГлу +NH3.
Освободившийся в канальцах почек NH3нейтрализуется собразованием солей аммония, которые выводятся с мочей:
NH3+ Н++ Сl-NH4Cl.
4)Синтез мочевины- это основной путь обезвреживания аммиака. На долю мочевины приходится 80% экскретируемого азота.
Процесс образования мочевины происходит в печени и представляет собой циклический процесс, который называется «орнитиновый цикл»(цикл Кребса–Гензелайта).
В цикле принимают участие две аминокислоты, которые не входят в состав белков – орнитин и цитруллин, и две протеиногенные аминокислоты – аргинин, аспарагин.
Процесс включает пять реакций: первые две протекают в митохондриях, остальные - в цитозоле гепатоцитов. Некоторые ферменты мочевинообразования есть в мозге, эритроцитах, сердечной мышце, однако весь набор ензимов есть только в печени.
І реакция– это синтез карбамоилфосфата:
СО2+NH3 + 2АТФNH2–CО–Ф + 2АДФ + Фн.
Фермент - карбамоилфосфатсинтаза І (митохонд–риальный). Существует также карбамоилфосфатсинтазаІІ(в цитозоле), которая участвует в синтезе пиримидиновых нуклеотидов.
Карбамоилфосфатсинтаза І - регуляторный фермент, для которого активатором является N–ацетилглутамат.
ІІ реакция– включение карбамоилфосфата в циклический процесс. В этой реакции происходит его конденсация с орнитином), в результате чего образуется цитруллин (реакция также происходит в митохондриях).
III реакция- образование аргининосукцината. Это вторая реакция, в которой используется энергия АТФ.
IV реакция - расщепление аргининосукцината с образованием аргинина и фумарата. Последний может поступать в ЦЛК, усиливая его работу. Т.о. это анаплеротическая (пополняющая) реакция для ЦЛК.
V реакция - регенерация орнитина с образованием мочевины.
Схема синтеза мочевины
С О2+NH3+ 2АТФкарбамоилфосфат + 2АДФ + Фн
О2+NH3+ 2АТФкарбамоилфосфат + 2АДФ + Фн
1
 NH2–CО–NH2
NH2–CО–NH2


 (мочевина) Орнитин
(мочевина) Орнитин
 Фн
Фн
52


АргининЦитруллин


 4
3АТФ
4
3АТФ


 Фумарат АМФ
Фумарат АМФ
АргининосукцинатФФн
Ферменты:
1 - карбамоилфосфатсинтаза;
2 - орнитинкарбамоилтрансфераза;
3 - аргининосукцинатсинтаза;
4 - аргининосукцинатлиаза;
5 - аргиназа (сильными ингибиторами фермента являются орнитин и лизин, конкурирующие с аргинином, активаторы - Са2+и Мn2+).
Орнитин, который восстанавливается в ходе цикла, может запускать новый цикл мочевинообразования. По своей роли орнитин аналогичен оксалоацетату в ЦЛК. Для прохождения одного цикла необходимо 3 АТФ, которые используются в 1–й и 3–й реакциях.
Орнитиновый цикл тесно взаимосвязан с ЦЛК.
Схематически взаимосвязь можно представить так:

 2 АТФ
2 АТФ

 Орнити-СО2
Орнити-СО2

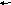 новый ЦЛК
новый ЦЛК
цикл
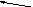
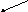 ОА
ОА

 Фумарат
АТФ
Фумарат
АТФ

Аспартат

Это «двухколесный велосипед» Кребса – ни одно колесо не способно «вращаться» без исправного функционирования второго.
Экскреция синтезированной мочевины обеспечивается почками. За сутки выделяется 20-35 г мочевины. При изменении количества белка в пище с целью поддержания азотистого равновесия скорость синтеза мочевины в организме изменяется:
белка с пищейсинтез ферментов цикласинтез мочевины,
если катаболизм белковсинтез мочевиныколичество
выводимогоазота.
Усиление катаболизма белков и, следовательно, повыше–ние экскреции мочевины наблюдаются при голодании и сахарном диабете.
При заболеваниях печени, которые сопровождаются нарушением синтеза мочевины, увеличивается концентра–ция аммиака в крови (гипераммониемия) и, как следствие, развивается печеночная кома.