

Государственное бюджетное образовательное учреждение высшего профессионального образования «Башкирский государственный медицинский университет» Министерства здравоохранения Российской Федерации
Лекции по дисциплине С.2.В.03. – Эмбриональный гистогенез и органогенез в норме и патологии
У ф а 2013
Составители: профессор, д.м.н. Х.Х. Мурзабаев, профессор, д.м.н. Ф.А. Каюмов.
Лекция № 1: Предмет, история развития и методы исследования в эмбриологии.
Название эмбриология происходит от греческих слов эмбрион - зародыш и логос - учение. Это название не соответствует содержанию современной науки. Эмбриология действительно описывает и выясняет всех процессов зародышевого развития - от оплодотворения яйца сперматозоидом до вылупления зародыша из яйцевых оболочек у яйцекладущих животных или до выхода его из материнского организма у живородящих. Однако эмбриология изучает и предзародышевый период - формирование половых клеток. Эмбриология изучает и так называемый постэмбриональный период. У млекопитающих животных некоторые системы органов (например, половая система, эндокринные железы) приобретают дефинитивные, т.е. окончательные, свойственные взрослому состоянию, структуры и функции спустя тот или иной промежуток времени после рождения. Зародыши многих животных, освобождаясь от яйцевых оболочек, имеют строение, мало сходное со строением взрослых организмов; у них развиваются провизорные (временные) органы, обеспечивающие их самостоятельное существование. Такие зародыши и личинки живут в совершенно отличной от имаго внешней среде и имеют специальные приспособления к этой среде. Впоследствии совершается метаморфоз, в ходе которого личиночные органы преобразуются, прежде чем достигнут своего дефинитивного состояния.
Таким образом, эмбриология занимается изучением индивидуального развития организма. Предметом ее исследований являются и регенерация, и бесполое размножение. Эмбриология изучает и патологические явления - причины нарушения нормального эмбрионального развития, возникновения уродств, причины нарушения нормальных процессов развития и жизни тканей и органов, Некоторые эмбриологические школы исследуют в своем аспекте причины возникновения опухолей.
Краткая история эмбриологии
Зачатки эмбриологических знаний о млекопитающих и птицах были уже в древнем Египте, Вавилонии, Ассирии, Индии и Китае.
Первые регулярные знания в области эмбриологии связывают с именем Гиппократа (460 - 370 до н.э.). Гиппократ предвосхитил идею преформации: «Все части зародыша образуются в одно и то же время. Согласно этой теории, каждый зародыш является уже вполне сформированным, имеющим все части организма, которому остается только расти. Выражаясь современным языком, в зародыше преобразованы и преформированы все особенности будущего организма, происходит лишь рост без дифференциации. Наиболее крайние преформисты представляли себе, что каждый организм, в том числе и человеческий, заключает в себе громадное количество поколений зародышей, вложенных друг в друга в теле прародителей со времени сотворения мира. Эта идея
господствовало в течение 17 - 18 веков - теория преформизма.
Рис. 1. Гомункулус – миниатюрный индивидуум, расположенный в головке сперматозоида, который использует яйцеклетку для питания в процессе своего роста.
 Между
представителями двух направлений
преформистов велись горячие споры.
Анималькулисты, или сперматики, например
А.В. Левенгук, описывал “семенных
зверушек” (сперматозоидов) использующих
запасы питательных веществ яйцеклетки
для своего роста. Овисты думали, что
зародыши в миниатюрном виде находятся
не в мужском семени, а в яйцеклетке, и
семя, входящее в яйцо при оплодотворении
составляет питательный материал
зародыша. Сторонниками преформизма
были выдающиеся ученые ХVII
и ХVIII
вв. А. Левенгук, Я. Сваммердам, М. Мальпиги,
А. Галлер, Ш. Бонне.
Между
представителями двух направлений
преформистов велись горячие споры.
Анималькулисты, или сперматики, например
А.В. Левенгук, описывал “семенных
зверушек” (сперматозоидов) использующих
запасы питательных веществ яйцеклетки
для своего роста. Овисты думали, что
зародыши в миниатюрном виде находятся
не в мужском семени, а в яйцеклетке, и
семя, входящее в яйцо при оплодотворении
составляет питательный материал
зародыша. Сторонниками преформизма
были выдающиеся ученые ХVII
и ХVIII
вв. А. Левенгук, Я. Сваммердам, М. Мальпиги,
А. Галлер, Ш. Бонне.
В 1V в. до н.э. работал другой величайший ученый античной древности -Аристотель (384 - 322 г.г. до н.э.). Аристотель сформулировал впервые теорию эпигенеза, значительно более соответствующую современной эмбриологии, Он, однако, внес в нее идеалистическое содержание.
Важнейшей вехой в истории эмбриологии считается 1759 г. В этот год была опубликована диссертация «Теория развития» двадцатишестилетнего Каспара Фридриха Вольфа, ставшего впоследствии академиком Петербургской Академии наук. В диссертации Фридрих Вольф впервые попытался объяснить возникновение новых растительных клеток при росте. Считал, что из уже имеющихся клеток-мешочков выдавливается жидкое вещество в виде капельки, поверхность капли затвердевает и капля превращается в новую клетку.
Вольф обосновал эпигенез, проследил развитие зародыша курицы, опровергнув преформизм. Большая заслуга Вольфа состояла в том, что он показал всю несостоятельность и абсурдность представлений преформистов о наличии в половой клетке готового организма, показав, что органы в эмбриогенезе возникают заново.
Весь XVI11 век прошел под знаком борьбы двух теорий развития. Явное торжество преформистских представлений тормозило разработку того прогрессивного начало, которое было заложено в теории эпигенеза. Накапливаемый фактический материал не получил должного признания: слишком низким был теоретический уровень науки.
Описание полного индивидуального развития - онтогенеза организма, начиная с яйцеклетки, впервые дал Карл Бэр (1792 - 1876). Он продолжил работу Вольфа на цыпленке и на основании полученных им фактов подтвердил некоторые выводы своего предшественника.
 Исследования
Бэра привели его к выводу, что развитие
состоит в постепенном усложнении более
простых структур.
Исследования
Бэра привели его к выводу, что развитие
состоит в постепенном усложнении более
простых структур.
Большая заслуга Бэра - открытие им яйца млекопитающих и человека. До него за яйцеклетку принимали так называемый графов пузырек - довольно значительных размеров образование, наполненное жидкостью, в стенке которого и находится яйцо.
Сравнивая развитие некоторых позвоночных, Бэр обратил внимание на то, что их зародыши обнаруживают между собой большее сходство, чем взрослые животные. При этом он отметил, что, чем моложе сравниваемые эмбриональные стадии, тем значительнее сходство. Закономерность, открытая Бэром, известна как явление зародышевого сходства.
Своим возникновением и развитием современная эволюционная эмбриология связана работами великих русских ученых А.О. Ковалевского (1840 - 1901) и И.И. Мечникова (1845- 1916).
Труды Ковалевского имели решающее значение для установления родственных связей между некоторыми группами животных. В этом отношении особенно большую роль сыграли его работы по изучению ланцетника и оболочников. Исследовав ранние стадии развития этих животных, А.О. Ковалевский доказал родство их с позвоночными и принадлежность к одному типу хордовых. Факты, добытые ученым, впервые наметили прямую связь между беспозвоночными, разделенными до того, казалось, непреодолимой пропастью.
Изучая эмбриональные стадии позвоночных и особенно мало изученных беспозвоночных, И.И. Мечников и А.О. Ковалевский показали, что развитие почти всех многоклеточных происходит через стадию образования трех зародышевых листков. Последние у животных сходны не только по способу происхождения, но также и по тем производным, которые дает каждый из них.
В четкой форме вопрос о связи эмбрионального развития с эволюцией впервые поставил Ф. Мюллер. Он сделал вывод о том, что в эмбриональном развитии наблюдается краткое повторение длительного исторического развития. Эта мысль была целиком принята Э. Геккелем и, подтвержденная новыми данными, получила более широкое обобщение в основном биогенетическом законе. Этот закон в наиболее общей формулировке гласит, что в своем индивидуальном развитии (онтогенезе) организм повторяет в краткой, сжатой форме историю своего вида (филогенез).
Экспериментальная эмбриология
Вильгельму Ру принадлежит честь основания экспериментального направления в эмбриологии, Он разрушал раскаленной иглой один из первых двух бластомеров лягушки. Из оставшегося бластомера развивалась половина зародыша. Такое же частичное развитие было обнаружено и в опытах на дробящихся яйцах некоторых других животных. Дефектные зародыши наблюдались при изоляции бластомеров асцидии, маллюсков, лошадиной аскариды, гребневиков и др.
Нарушения в развитии при изоляции бластомеров или даже отдельных частей яйца Ру объяснял предопределением в яйце частей будущего организма. Яйцо представляло как бы мозаику из зачатков органов, изъятие части мозаики вызывало отсутствие определенных органов. Одновременно с ним некоторые другие ученые стали использовать различные экспериментальные методики в своих исследованиях. Вскоре начали экспериментировать Г. Дриш, Ж. Леб и многие другие. Г. Дриш, которому эмбриология обязана выдающимися экспериментами по изоляции бластомеров.
Для прослеживания судьбы тех или иных бластомеров, для изучения передвижения клеточного материала в ходе развития важное значение имела разработанная В. Фогтом методики меток, наносимых витальным красителем на отдельные части зародыша. Эта методика дала возможность выяснить процессы гаструляции у амфибий и других животных.
Исключительное значение, притом все возрастающее в наше время, имела разработка методик культивирования тканей и зачатков органов вне организма, соответствующие способы хирургии, набор питательных сред, способы их стерилизации. Однако честь открытия метода тканевых культур принадлежит Р. Г. Гаррисону.
Наибольшее влияние на экспериментальную эмбриологию в XX в. оказала школа Ганса Шпемана, предложившего свою теорию индивидуального развития и разработавшего прекрасные методики микрохирургии на зародышах: снятие оболочек яиц животных, пересадка частей одного зародыша другому, изготовление благоприятной жидкой среды для развития и др. Шпеману и его ученикам удалось установить взаимозависимость частей развивающегося зародыша.
Одна из самых плодотворных теорий развития, которая объединяет усилия эмбриологов на протяжении всего XX в. и по настоящее время, - теория эмбриональной индукции.
Экспериментальная разработка будущей теории началась с разнообразных опытов по пересадке закладок у ранних зародышей амфибий в лаборатории Ганса Шпемана.
Немецкий ученый Г. Шпеман впервые установил, что закладка нервной системы у земноводных связана с материалом хорды, который, перемещаясь внутрь зародыша, располагается под дорзальной эктодермой, развивающейся в нервную систему. Материал хорды, который определяет закладку центральной нервной системы, был назван Шпеманом организационным центром.
Наличие формообразовательных влияний было установлено и при развитии ряда других органов. Впервые это было показано на примере развития глаза. Оказалось, у большинства исследованных животных при удалении зачатка глаза до его контакта с покрывающей эктодермой хрусталик не развивается.
Формообразовательное влияние при развитии глаза не является односторонним. Хрусталик со своей стороны действует на мозг.
Взаимодействие частей зародыша, в результате которого определяется развитие органов, получило название индукции, а сами части, определяющие развитие, - индукторов.
Исключительная роль в развитии эволюционной эмбриологии принадлежит отечественным эмбриологам Д.П. Филатову и П.П. Иванову. Разработали свои методики микрохирургии и положили начало сравнительной экспериментальной эмбриологии.
Современная эмбриология ставит своей задачей управление развитием организмов, Осуществление этой задачи возможно при условии тесной связи эмбриологии с другими науками, в первую очередь с гистологией и цитологией. Эмбриология должна быть теснейшим образом связана с генетикой и цитогенетикой. Тесная связь эмбриологии с экологическими науками проявляется при изучении влияния внешней среды на развитие организмов.
Лекция № 2: Эмбриональное развитие сердечно-сосудистой системы, органов кроветворения и иммуногенеза. Аномалии развития.
Возникновение кровеносных сосудов тесно связано с возникновением крови. Источник развития у них общий - мезенхима.
Первые кровеносные сосуды возникают на 2-й неделе, вне тела эмбриона, в мезенхиме стенки желточного мешка в виде так называемых кровяных островков. Клетки, расположенные по периферии этих островков, - ангиобласты - митотически активно размножаются. Эти клетки уплощаются, вступают в связи друг с другом, образуя стенку сосуда. Клетки центральной части островка округляются и превращаются в клетки крови.
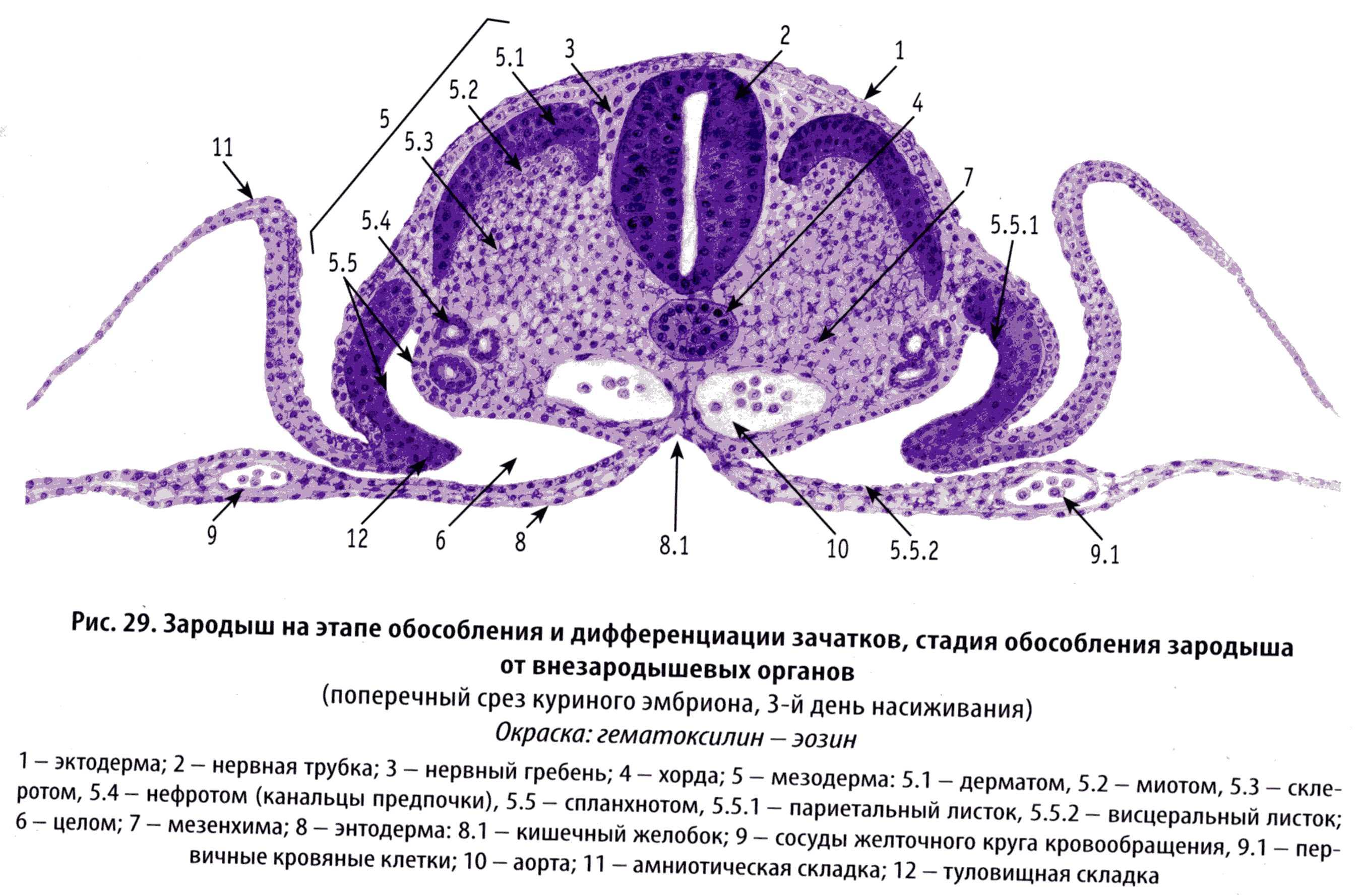
В теле зародыша из мезенхимы образуются первичные кровеносные сосуды, имеющие вид трубочек и щелевых пространств, но без клеток крови внутри. В конце 3-й недели внутриутробного развития сосуды тела зародыша сообщаться с сосудами внезародышевых органов.
 Дальнейшее
развитие сосудов происходит после
начала циркуляции крови под влиянием
тех гемодинамических условий (кровяное
давление, скорость кровотока), которые
создаются в различных частях тела, что
обусловливает появление специфических
особенностей строения стенки внутриорганных
и внеорганных сосудов. Из мезенхимных
клеток, окружающих сосуд, позднее
дифференцируются гладкие мышечные
клетки, перициты и а адвентициальные
клетки, а также фибробласты.
Дальнейшее
развитие сосудов происходит после
начала циркуляции крови под влиянием
тех гемодинамических условий (кровяное
давление, скорость кровотока), которые
создаются в различных частях тела, что
обусловливает появление специфических
особенностей строения стенки внутриорганных
и внеорганных сосудов. Из мезенхимных
клеток, окружающих сосуд, позднее
дифференцируются гладкие мышечные
клетки, перициты и а адвентициальные
клетки, а также фибробласты.
 В
эмбриогенезе человека сердце закладывается
очень рано, когда зародыш еще не обособлен
от желточного мешка и кишечная энтодерма
одновременно представляет собой
внутреннюю выстилку последнего. В это
время в кардиогенной зоне в шейной
области между энтодермой и висцеральными
листками спланхнотомов слева и справа
скапливаются клетки мезенхимы, образующие
и справа, и слева клеточные тяжи. Эти
тяжи вскоре превращаются в эндотелиальные
трубки. В дальнейшем мезенхимные трубки
сливаются и из их стенок образуется
эндокард.
Сразу
же нужно отметить, что закладки эндокарда
и сосудов в принципе тождественны. Та
область висцеральных листков спланхнотомов,
которая прилежит к этим трубкам, получила
название миоэпикардиальных
пластинок. Из
этих пластинок дифференцируются две
части: одна - внутренняя, прилежащая к
мезенхимной трубке, превращается в
зачаток миокарда,
а
из наружной образуется эпикард
(рис.
2). Первоначально сердце представляет
собой прямую трубку, в которой различают:
В
эмбриогенезе человека сердце закладывается
очень рано, когда зародыш еще не обособлен
от желточного мешка и кишечная энтодерма
одновременно представляет собой
внутреннюю выстилку последнего. В это
время в кардиогенной зоне в шейной
области между энтодермой и висцеральными
листками спланхнотомов слева и справа
скапливаются клетки мезенхимы, образующие
и справа, и слева клеточные тяжи. Эти
тяжи вскоре превращаются в эндотелиальные
трубки. В дальнейшем мезенхимные трубки
сливаются и из их стенок образуется
эндокард.
Сразу
же нужно отметить, что закладки эндокарда
и сосудов в принципе тождественны. Та
область висцеральных листков спланхнотомов,
которая прилежит к этим трубкам, получила
название миоэпикардиальных
пластинок. Из
этих пластинок дифференцируются две
части: одна - внутренняя, прилежащая к
мезенхимной трубке, превращается в
зачаток миокарда,
а
из наружной образуется эпикард
(рис.
2). Первоначально сердце представляет
собой прямую трубку, в которой различают:
Верхний конец – луковица, переходящая в артериальный конус.
Средний отдел – собственно сердце.
Нижний отдел – венозный синус.
Уже в эти сроки сердце начинает пульсировать и обуславливает циркуляцию кровяных телец.
Одном из основных факторов, характеризующих ранние этапы развития сердца, является быстрый рост в длину первичной сердечной трубки, которая увеличивается в длину быстрее, чем полость, в которой она расположена (полость перикарда). Это обстоятельство является одной из причин того, что сердечная трубка, увеличиваясь в длину, образует петлю. Ее передняя нисходящая часть – общий желудочек, венозный конец загибается назад и вверх. При этом венозный отдел растет в краниальном направлении и охватывает сзади и с боков артериальный конус, а артериальный отдел сильно разрастается и смещается каудально. В результате в развивающемся сердце эмбриона можно видеть контуры его основных дефинитивных отделов - предсердий и желудочков.
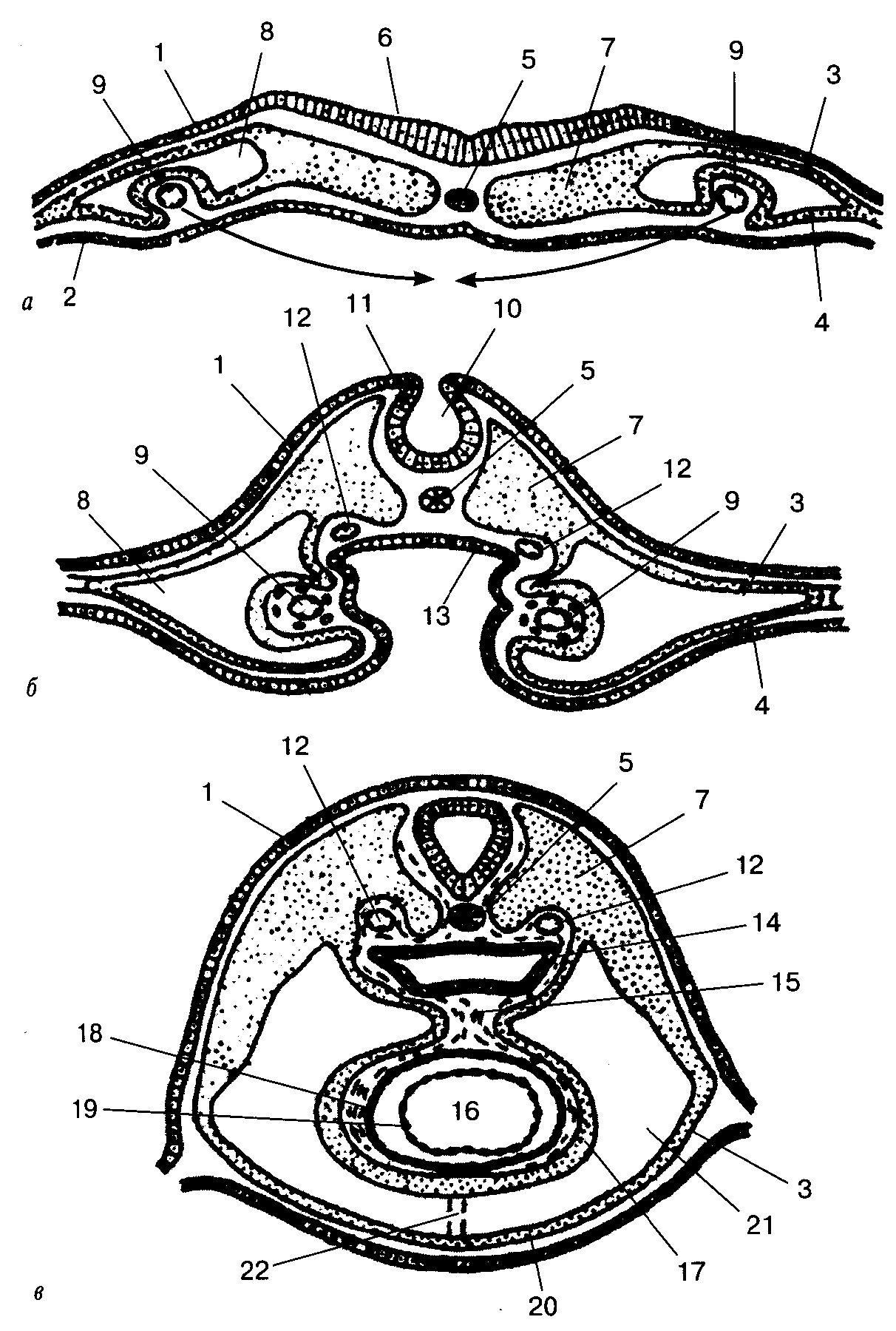
Дальнейшие изменения приводят к образованию четырехкамерного сердца (рис. 4). В начале венозный и артериальный отделы разделены поперечной перетяжкой. Сообщаются эти отделы через узкий ушковый канал. Двухкамерное сердце существует в эмбриогенезе человека недолго и преобразуется с возникновением продольных перегородок в четырехкамерное. Изменения, приводящие к возникновению четырехкамерного сердца и образованию основных структур, соответствующих картине дефинитивного сердца, заканчиваются в основном к концу третьего месяца эмбриональной жизни.
Развитие
закладки эндокарда, как указывалось,
принципиально соответствует процессам,
происходящим при дифференцировке
сосудистой стенки. К эндотелиальной
трубке, образовавшейся на самых ранних
этапах, присоединя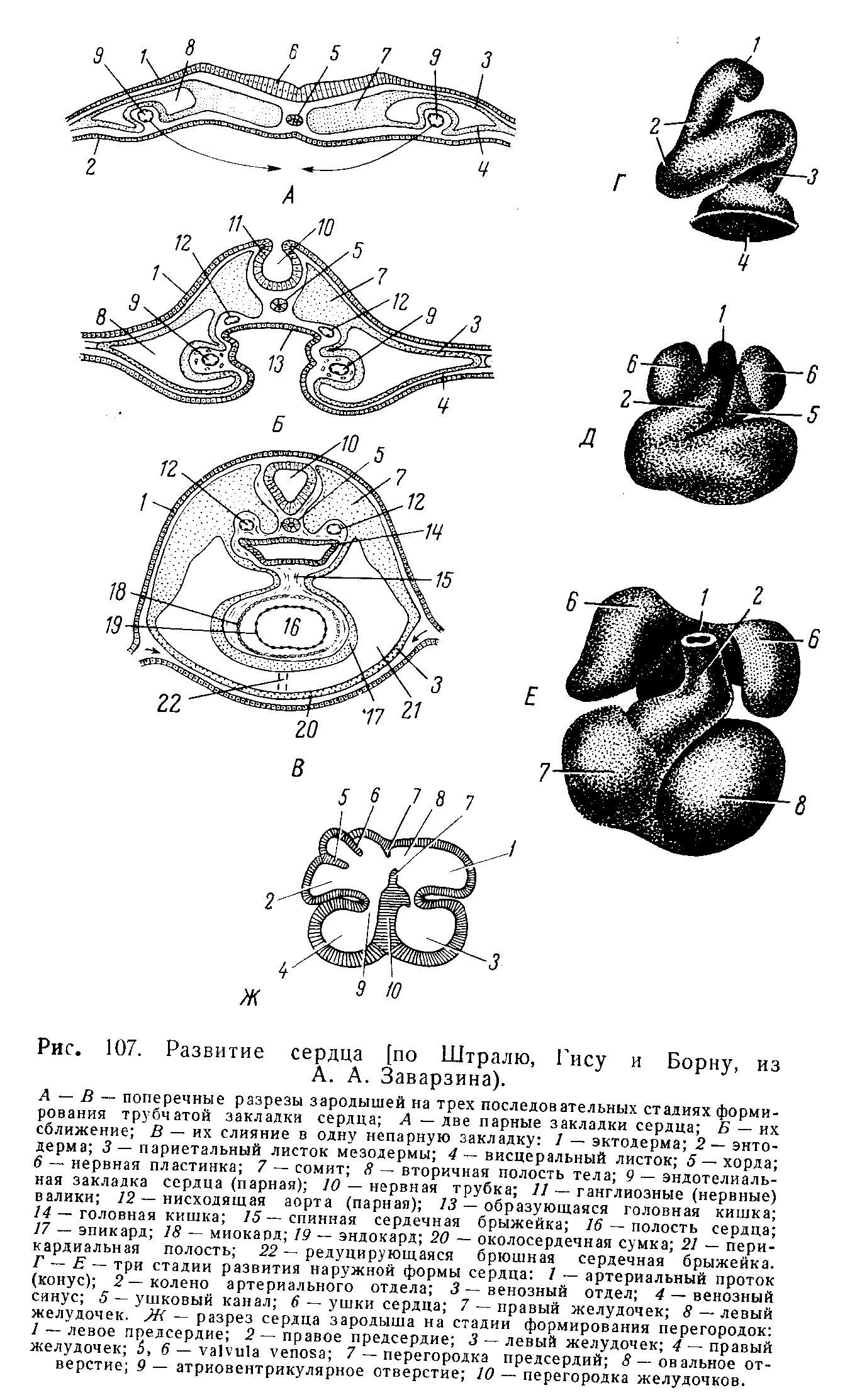 ются
впоследствии дифференцирующиеся из
окружающей мезенхимы субэндотелий,
эластический аппарат, коллагеновые
волокна и гладкие мышцы.
ются
впоследствии дифференцирующиеся из
окружающей мезенхимы субэндотелий,
эластический аппарат, коллагеновые
волокна и гладкие мышцы.
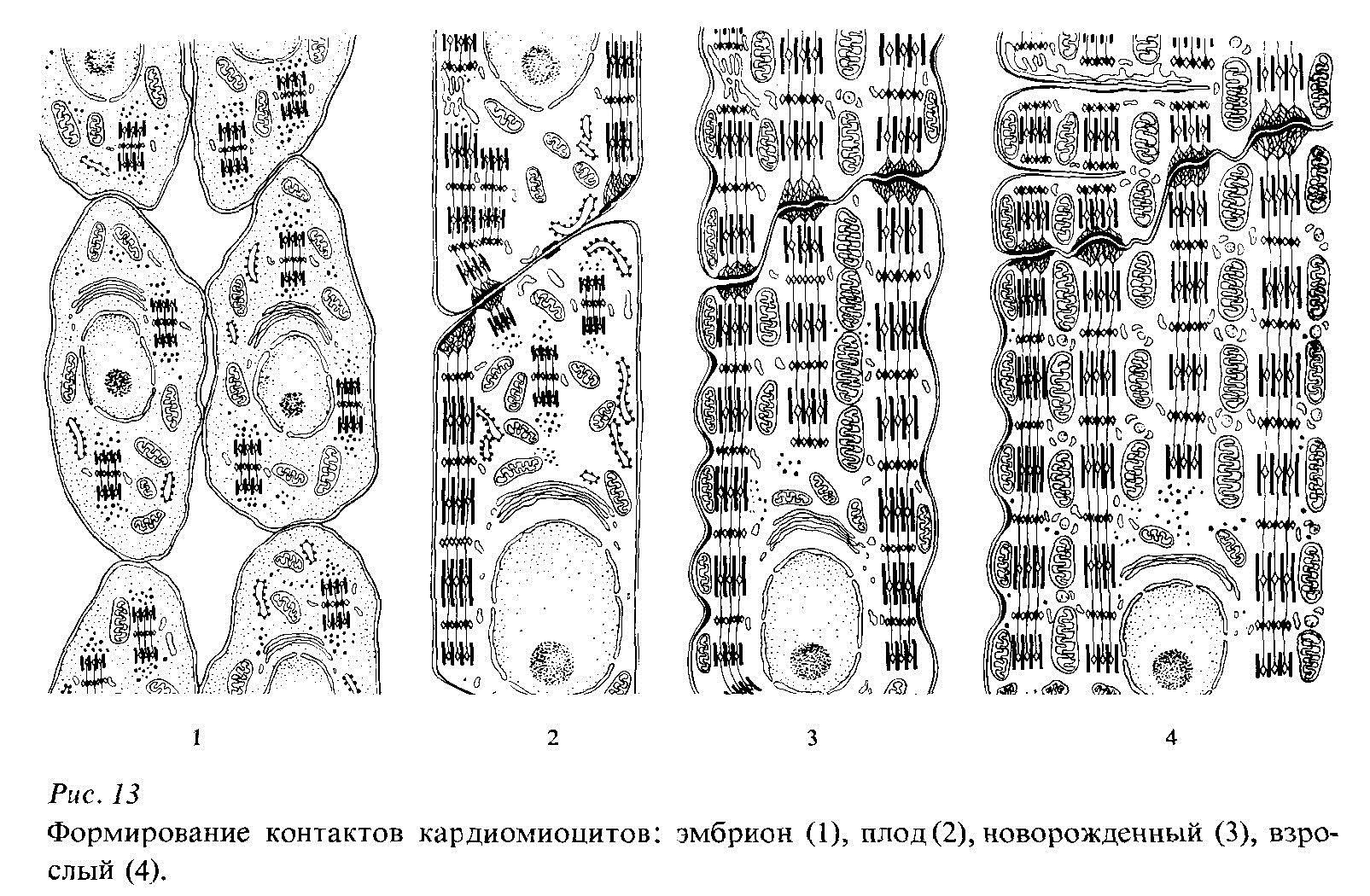
В миоэпикардиальной пластинке также заметны процессы дифференцировки. Прежде всего, на наружной ее поверхности, обращенной к целомической полости, возникает эпителиоподобный слой клеток с расположенной под ним соединительной тканью. Иными словами, возникает закладка эпикарда. Только после этого активизируются гистогенетические процессы, приводящие к формированию миокарда. Клетки миокарда - кардиомиобласты - лежат первоначально рыхло, на довольно значительном расстоянии друг от друга (рис. 5). В дальнейшем миобласты устанавливают контакт друг с другом. В местах контакта их мембраны представляются утолщенными в определенных зонах за счет накопления электронноплотных гранул. Такие гранулы, находящиеся вне связи с фибриллярным материалом, образуют типичные десмосомы. В периферических частях цитоплазмы клеток миокарда появляются первые тонкие миофиламенты, сгруппированные в рыхлые пучки Гранулы, связанные с пучками миофиламентов, рассматриваются как примитивные вставочные пластинки. Ранние вставочные пластинки могут проходить косо по отношению к оси волокна. Однако постепенно каждый диск ориентируется под прямым углом к оси волокна (миофибриллам). Такая структура вставочных дисков характерна для новорожденного.
Увеличение массы миокарда в эмбриональном периоде происходит как за счет митозов, так и за счет увеличения размеров клеток. Увеличение диаметра волокон миокарда связано с увеличением массы цитоплазмы, главным образом, за счет новообразования миофибрилл внутри каждой клетки. Постепенно в дифференцирующихся мышечных клетках увеличивается количество митохондрий. Митохондрии постепенно удлиняются и располагаются упорядоченно между миофибриллами параллельно их длине.
Вообще же постоянно меняющиеся гемодинамические условия в растущем организме приводят к соответствующим изменениям гистоструктур сердца, в том числе и миокарда. В связи с этим становление дефинитивных структур сердца занимает длительный период онтогенеза, включающий многие годы постнатального периода.
Миокард
является многотканевой структурой. В
его построении участвуют не только
мышечная, но и соединительная ткань.
Миокард эмбриона содержит небольшое
количество соединительной ткани.
Коллагеновые волокна отмеча
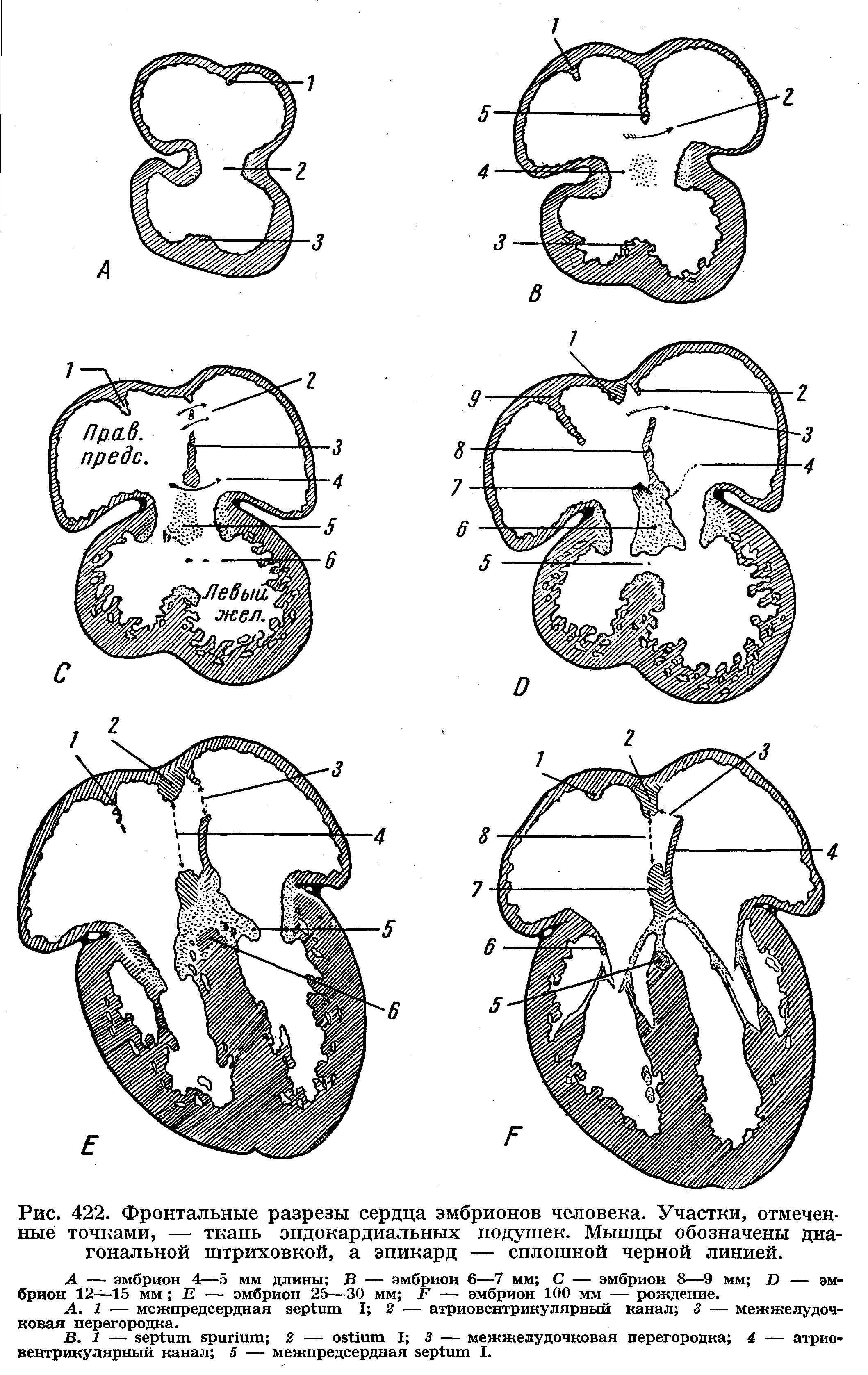 ются
лишь вблизи сосудов. Эластических
волокон очень мало. Сосудистая система
сердца эмбриона относится к так
называемому рассыпному типу.
ются
лишь вблизи сосудов. Эластических
волокон очень мало. Сосудистая система
сердца эмбриона относится к так
называемому рассыпному типу.
Имеется множество описаний нервных элементов (клеток, волокон) в стенке сердца эмбрионов различных возрастов. На человеческом материале показано наличие нейробластов в стенке 7-недельного эмбриона. Развитие нейронов протекает неравномерно и характеризуется волнообразностью. К моменту рождения дифференцировка интрамуральных нейронов не завершена: они находятся на разных стадиях развития, причем зрелые нейроны - единичны.
 Стенка
сердца новорожденного тонкая, легко
растяжимая. Эндокард представлен слоем
эндотелия, субэндотелием. Гладкомышечные
клетки, как правило, единичные: мышечный
слой эндокарда формируется позже.
Волокна миокарда тонкие, составлены
мелкими клетками. Соединительнотканная
строма, жировая ткань развиты очень
слабо. Внешняя форма сердца округлая с
большим поперечным диаметром. Верхушка
его почти всегда образована правым
желудочком. Относительный вес сердца
велик: у новорожденных он составляет
примерно 0,8% веса тела.
Стенка
сердца новорожденного тонкая, легко
растяжимая. Эндокард представлен слоем
эндотелия, субэндотелием. Гладкомышечные
клетки, как правило, единичные: мышечный
слой эндокарда формируется позже.
Волокна миокарда тонкие, составлены
мелкими клетками. Соединительнотканная
строма, жировая ткань развиты очень
слабо. Внешняя форма сердца округлая с
большим поперечным диаметром. Верхушка
его почти всегда образована правым
желудочком. Относительный вес сердца
велик: у новорожденных он составляет
примерно 0,8% веса тела.
После рождения проходит длительный период времени, пока структура сердца не достигнет дефинитивного состояния. В это время имеет место увеличение массы органа и значительные изменения его внутренней структуры. Такая динамика структур сердца связана со значительными изменениями гемодинамики, связанными в свою очередь с множеством факторов: выключением плацентарного кровообращения, началом функционирования малого круга кровообращения, ростом и дифференцировкой органов и тканей и т. д.
Аномалии развития сердца
Акардия – отсутствие сердца (нежизнеспособный плод).
Дистопия сердца – чаще всего декстракардия – правостороннее положение сердца.
Дефект межпредсердной перегородки.
Дефект межжелудочковой перегородки:
- отсутствие перегородки (трехкамерное сердце);
- межжелудочковое отверстие.
5. Пороки предсердно-желудочковых клапанов, клапанов аорты и легочного ствола.
6. Общий артериальный ствол – неразделенные легочной ствол и восходящая аорта, приводит к смешению артериальной и венозной крови.
7. Открытый артериальный (Боталлов) проток – наличие соединения между дугой аорты и левой легочной артерией.
8. Триада Фалло:
- дефек межжелудочковой перегородки;
- стеноз легочного ствола;
- гипертрофия миокарда правого желудочка.
9. Тетрадо Фалло (40-50% всех пороков сердца) – к триаде Фалло присоединяется декстрапозиция аорты.
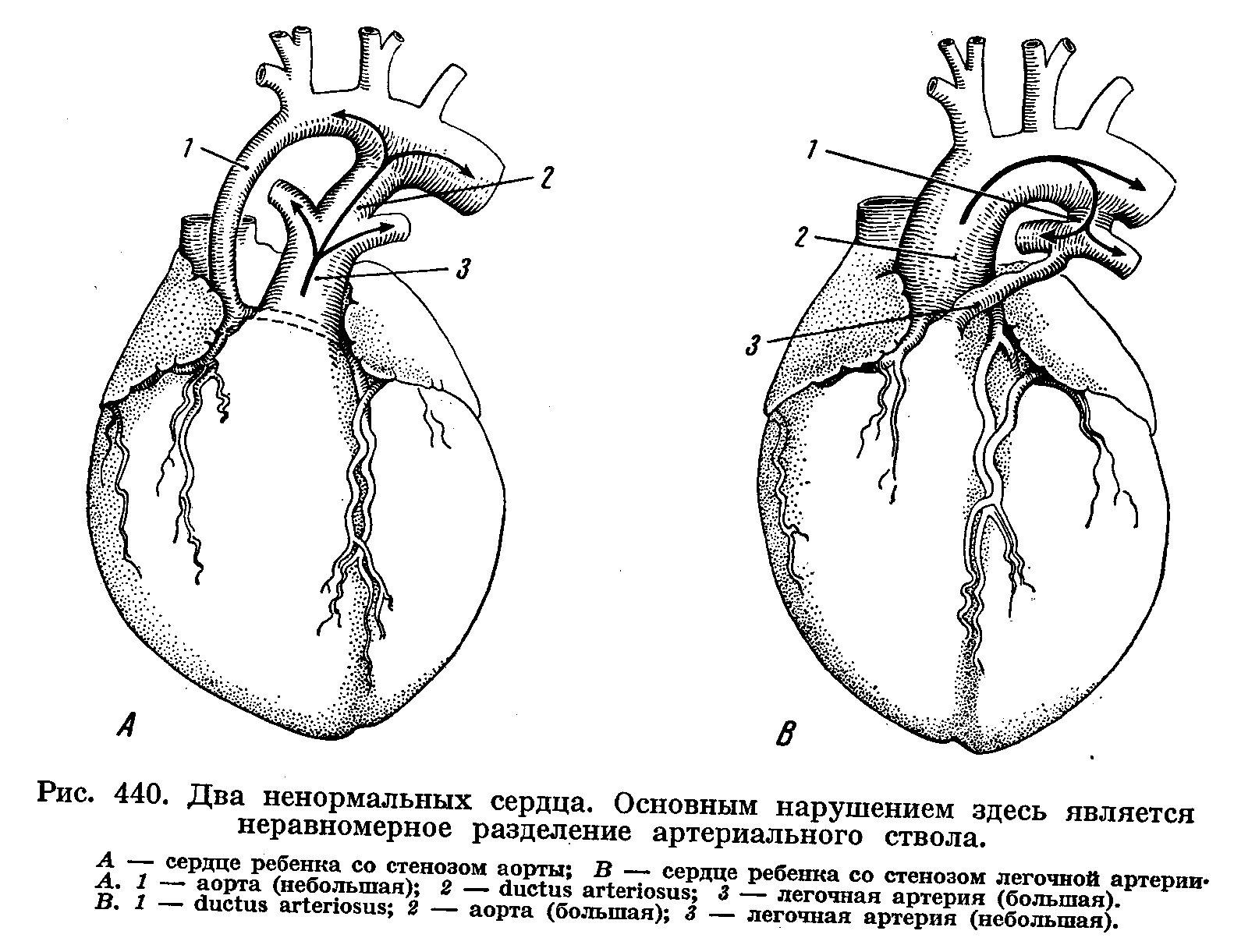
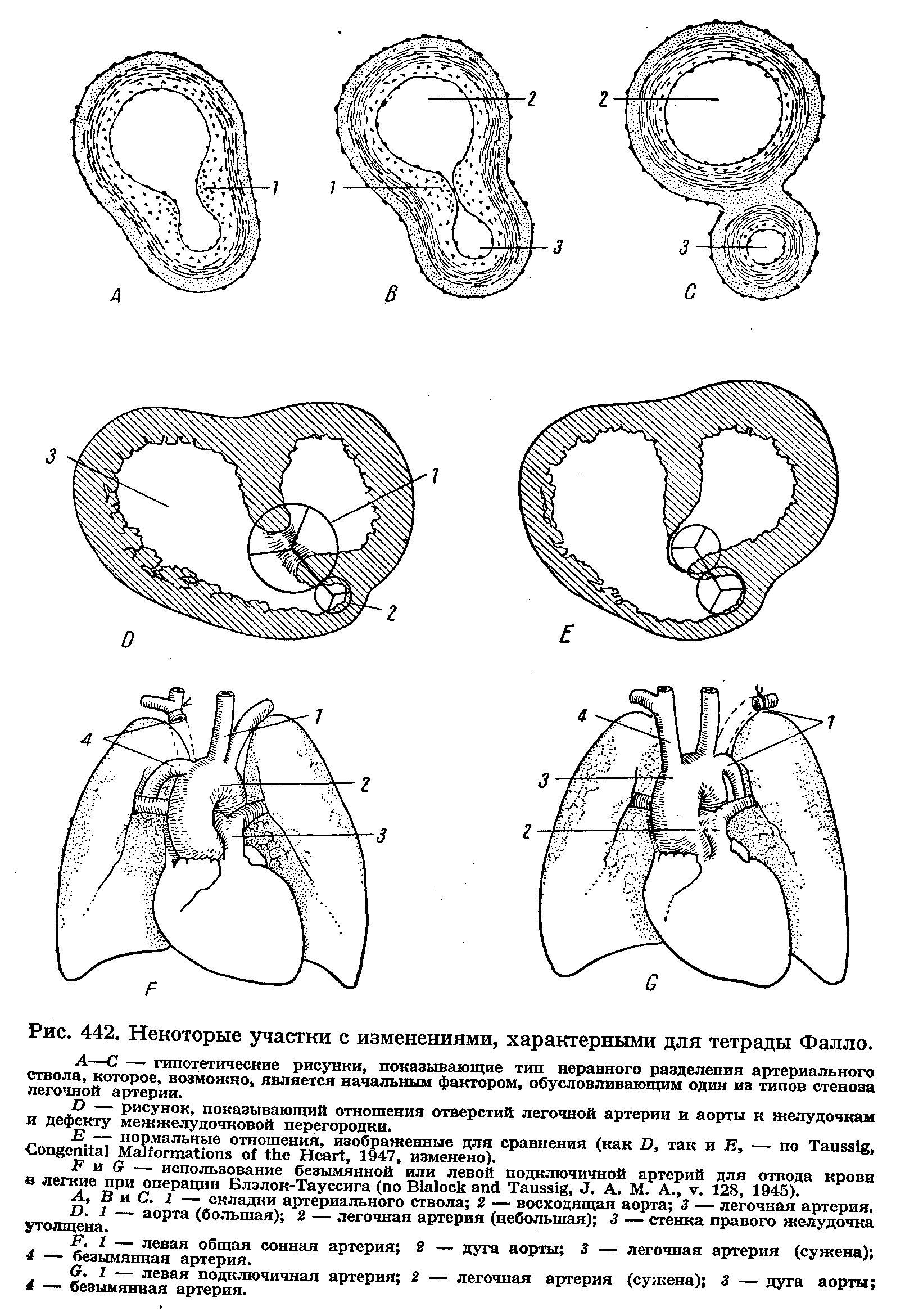

Рис. Дефекты межпредсердной перегородки
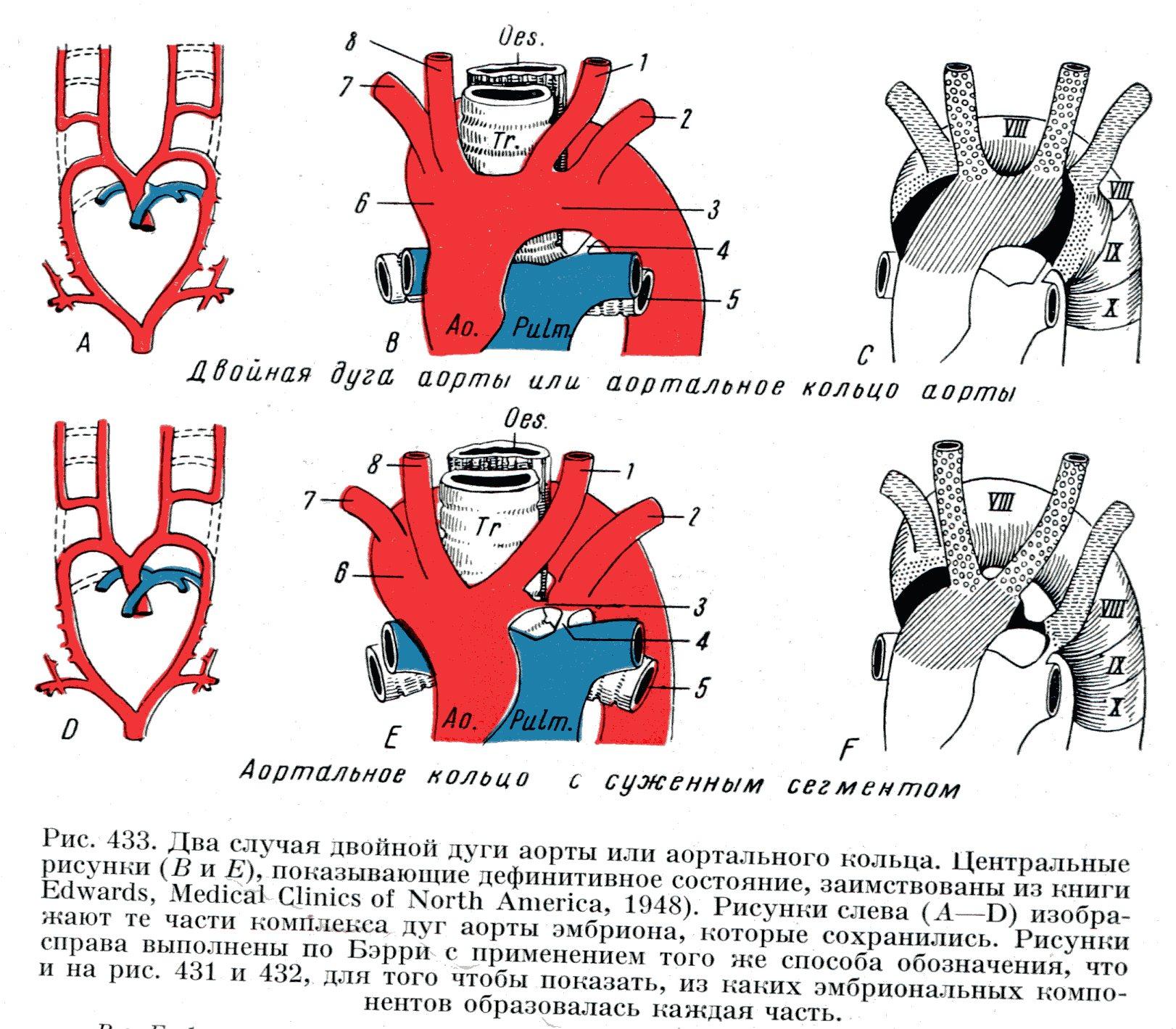
У эмбриона артериальный конус разделяется на 2 вентральные аорты, идущие в краниальном направлении, а в области шеи они поворачивают назад и вниз, и распадаясь на 6 пар аортальных дуг проходят через соответствующие жаберные дуги и сливаясь продолжаются в 2 дорсальные аорты. Дорсальные аорты сливаются в непарную нисходящую аорту, которая разделяется в нижней части туловища на 2 пупочные артерии, идущие в аллантоис, позднее в плаценту.

При редукции жаберного аппарата происходит трансформация вышеописанных сосудов:
Артериальный конус разделяется перегородкой на легочной ствол (передняя часть) и восходящую аорту;
I, II и V пары аортальных дуг редуцируются;
III пара аортальных дуг преобразуются во внутренние сонные артерии;
IV пара аортальных дуг: левая – превращается в дугу аорты, правая – в плечеголовной ствол и в правую подключичную артерию;
VI пара аортальных дуг – в легочные артерии: правая дуга – правая легочная артерия, левая дуга – левая легочная артерия и артериальный проток.
Аномалии развития артерий
Стеноз (сужение) или каорктация (перетяжка) аорты и легочного ствола, их ветвей.
Декстрапозиция дуги аорты.
Две дуги аорты (сохранение IV пары аортальных дуг).
Удвоение дуги аорты.
Аортальное кольцо – охватывает трахею и пищевод.
Аномалии ветвления дуги аорты - многочисленные вариации в превращениях первичных аортальных дуг. Они выражаются, в частности, в отхождении от дуги аорты не трех, как в норме, а четырех-пяти ветвей. Встречается и крайний случай, когда все ветви отходят в виде одного общего ствола.


4. Аномалии вен.
Примеры:
- впадение одной из легочных вен в верхнюю полую вену, левую безымянную или непарную вену;
- двойная верхняя полая вена;
- атипичное расположение нижней полой вены.
Органы кроветворения и иммуногенеза
Кровь во взрослом организме - одна из важнейших интегрирующих систем. И в процессе эмбриогенеза она является носительницей факторов, регулирующих процессы роста и дифференцировки органов и тканей. Потребность в такой системе, выполняющей трофические и координирующие функции, возникает очень рано. В соответствии с этим развитие крови (и связанный с ним процесс образования сосудов) начинается очень рано. Так, первые признаки начинающегося развития первичных сосудов и крови у человеческого эмбриона отмечены уже в конце второй недели развития.
Становление кроветворения в онтогенезе человека проходит ряд этапов не только количественными и качественными параметрами кроветворного процесса, но и его локализацией.
В течение внутриутробного развития место образования форменных элементов крови (гемопоэз) несколько раз меняется. Наиболее ранним из них служит желточный мешок, а позднее его сменяют печень и селезенка, а далее - костный мозг и лимфоидные органы.
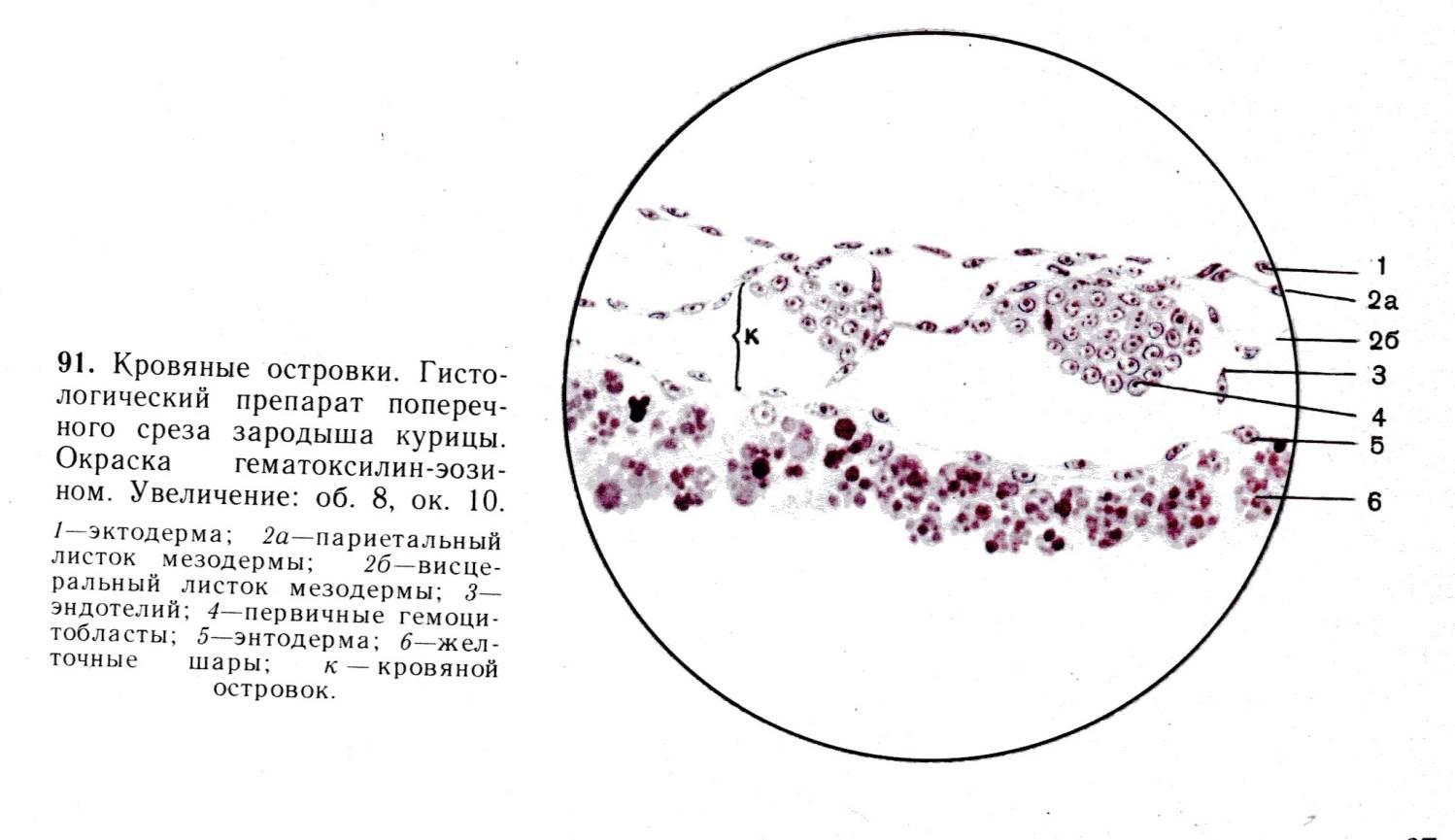
Кроветворение в стенке желточного мешка (2 – 10 нед.) является внезародышевым, поскольку он относится к внезародышевым провизорным органам. Оно тесно связано с развитием первых сосудов, которые появляются в мезенхиме стенки желточного мешка с возникновением кровяных островков на 2-й неделе эмбрионального развития.
Кровяные островки образуются в мезенхиме вследствие индуцирующего влияния энтодермы желточного мешка. Они имеют вид мелких компактных скоплений округлившихся мезенхимных клеток. Расположенные в центральной части островка клетки превращающихся в стволовые клетки крови (СКК). Периферические клетки кровяных островков (ангиобласты) уплощаются и образуют соединения между клетками, формируют эндотелиальную выстилку. Сливаясь друг с другом, кровяные островки образуют в стенке желточного мешка сосудистую сеть.
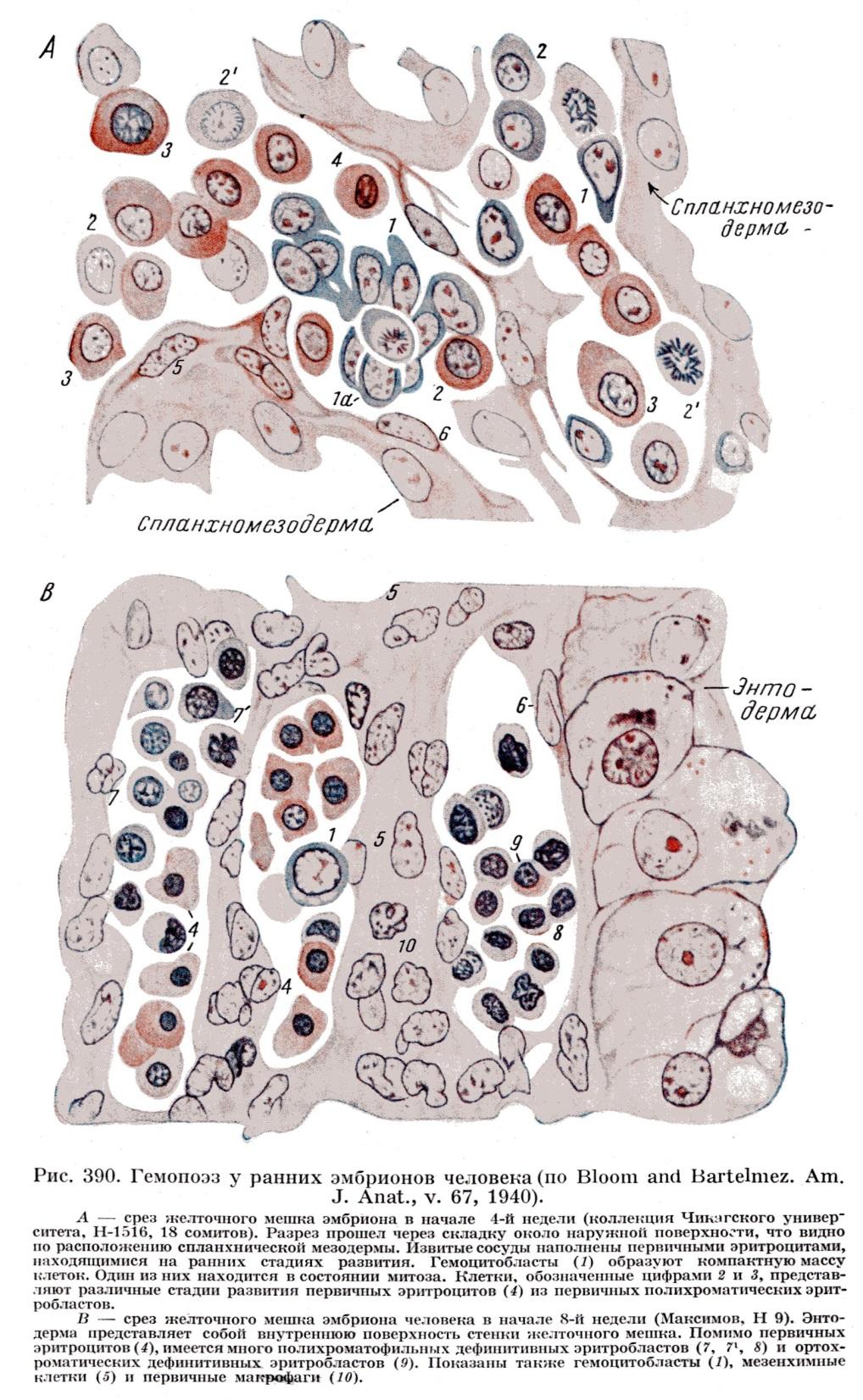
СКК в кровяном островке делятся и дифференцируются в первичные эритробласты (называемые из-за своих больших размеров также мегалобластами - от греч. megas -большой и blastos - росток). Мегалобласты - крупные клетки с базофильной цитоплазмой, которые по мере накопления гемоглобина (HbF - фетального) превращаются в полихроматофильные, а затем в оксифильные эритробласты (мегалоцитам). Описанное мегалобластическое кроветворение свойственно эмбриональному периоду, но может возникать после рождения при тяжелом заболевании крови - злокачественной анемии (обусловленной недостаточностью витамина В 12). Наряду с эритропоэзом, происходящим внутри сосудов (интраваскулярно), вне сосудов (экстраваскулярно) образуются в небольшом числе гранулоциты.
Из желточного мешка СКК мигрируют в печень, селезенку и другие кроветворные органы, где продолжаются следующие периоды гемопоэза.
Кроветворение в печени осуществляется, начиная с 5-6-й недели внутриутробного развития и достигает максимальной активности на 2-м месяце (когда кроветворение на 80% обеспечивается печенью и на 20% - селезенкой), стихает к концу внутриутробного развития и полностью завершается в течение первых двух недель после рождения. В печени из СКК, преимущественно экстраваскулярно, дифференцируются эритроциты, в меньшем количестве - гранулоциты и мегакариоциты.
Кроветворение в селезенке протекает экстраваскулярно и начинается позднее, чем в печени (с середины 3-го мес.), достигая наибольшей активности с 4-го по 6-й мес. Селезенка как орган эритро- и миелопоэза активно функционирует в эмбриональном периоде очень недолго.
3 – 7 месяцы эмбрионального развития в селезенке преобладает эритро- и гранулоцитопоэз, лимфоидные клетки располагаются мелкими, диффузно распределенными группами. К 8-му мес. внутриутробной жизни появляются лимфоидные скопления, группирующиеся преимущественно вокруг артерий, усиливается лимфоцитопоэз. К концу эмбрионального периода в селезенке эритро- и гранулоцитопоэз затухают, усиливается лимфопоэз.
При некоторых патологических изменениях системы крови (миелопролиферативных заболеваниях), когда красный костный мозг оказывается не в состоянии производить достаточное количество клеток крови, гемопоэз может вновь возникнуть в печени и селезенке (экстрамедуллярное кроветворение).
Кроветворение в тимусе начинается со 2-го мес. внутриутробного развития и протекает с образованием Т-лимфоцитов, которые в дальнейшем расселяются в лимфоидные органы - селезенку и лимфатические узлы (закладываются на 9-10-й нед.).
Кроветворение в костном мозге начинается с 3-го мес. внутриутробного развития. СКК заселяют полости в образующихся костях и дают начало всем видам форменных элементов крови. На 5-м мес, в нем образуются лейкоциты и тромбоциты., на 7-м - эритроциты. Костный мозг замещает печень и селезенку в качестве кроветворного органа и становится окончательным (дефинитивным) центральным органом гемопоэза в конце развития плода, оставаясь таковым у новорожденного, ребенка и взрослого.
Лекция № 3: Эмбриональный гистогенез и органогенез пищеварительной системы. Аномалии развития.
В своем развитии пищеварительная система проходит несколько этапов. На первом этапе происходит формирование кишечной трубки и параллельно с этим - процесс развития ротовой полости и заднего прохода; следующим этапом является образование закладок пищевода, желудка, тонкого и толстого кишечника с брыжейками; после чего из кишечной стенки формируются отдельные органы; и, наконец, следует этап образования тканей в каждом из развивающихся органов.

Образование пищеварительной системы начинается с появления энтодермы. Этот листок зародышевого щитка представляет собой вначале плоскую пластинку, которая позже прогибается и постепенно превращается в продольный желобок энтодермы, еще раскрытый со стороны полости желточного пузыря. Вследствие разрастания зародышевого диска происходит приподнимание его над желточным пузырем и зародыш остается соединенным с ним только посредством сравнительно узкого стебля. Уже у трехмиллиметрового зародыша (3 неделя развития) отчетливо различаются три части: передняя кишка, средняя кишка, соединенная с полостью желточного мешка, и задняя кишка, причем
передняя и задняя кишка заканчивается пока слепо. По мере удлинения зародыша весь зачаток кишечника все больше принимает характер трубки.
Энто дерма
очень рано оказывается тесно связанной
с висцеральным листком спланхнотомов.
В промежуток между этими листками
впоследствии врастает мезенхима.
Энтодерма дает эпителий кишечного
тракта и его желез, мезенхима -
соединительную ткань с сосудами и
мышечную оболочку, а висцеральный листок
спланхнотомов – наружную серозную
выстилку кишечника.
дерма
очень рано оказывается тесно связанной
с висцеральным листком спланхнотомов.
В промежуток между этими листками
впоследствии врастает мезенхима.
Энтодерма дает эпителий кишечного
тракта и его желез, мезенхима -
соединительную ткань с сосудами и
мышечную оболочку, а висцеральный листок
спланхнотомов – наружную серозную
выстилку кишечника.
Превращение кишечной трубки в сплошной канал происходит следующим образом. На головном конце зародыша образуется впячивание кожной эктодермы, которое растет навстречу слепому концу передней кишки. Образовавшаяся таким образом ротовая бухта, отделенная от передней кишки ротоглоточной мембраной, представляет собой зачаток первичной ротовой полости. В конце 3-й недели развития ротоглоточная мембрана прорывается и тогда устанавливается прямое сообщение между передней кишкой и внешней средой.
В каудальном конце зародыша образуется небольшое впячивание эктодермы, которое контактирует с каудальным концом слепой задней кишки, образуя двуслойную клоакальную мембрану, она прорывается и кишечник вступает в сообщение с полостью клоаки. Таким образом, кишечная трубка становится открытой с обеих сторон.