

Кинетика оксигенирования миоглобина и гемоглобина
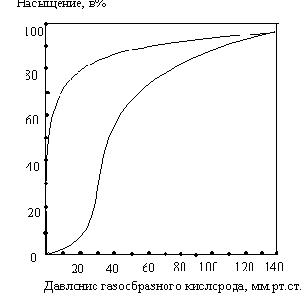
Почему миоглобин не способен транспортировать кислород, но зато активно его запасает? Для миоглобина изотерма адсорбции кислорода имеет форму гиперболы. Давление О2 в ткани, окружающей легочные капилляры, составляет 100 мм рт.ст., поэтому миоглобин в легких мог бы весьма эффективно насыщаться кислородом. В венозной крови Р О2 равно 40 мм рт. ст., а в активно работающей мышце - 20 мм рт.ст. Но даже при парциальном давлении 20 мм рт. ст. степень насыщения миоглобина кислородом будет весьма значительна, и поэтому миоглобин не может служить транспортной молекулой для доставки О2 от легких к периферическим тканям. Однако при кислородном голодании, которым сопровождается тяжелая физическая работа, парциальное давление О2 в тканях может понизиться и до 5 мм рт.ст.; при столь низком давлении миоглобин легко отдает кислород, обеспечивая тем самым окислительный синтез АТФ в митохондриях мышечных клеток.
Кинетика оксигенирования гемоглобина коренным образом отличается от кинетики оксигенирования миоглобина. Кривая насыщения гемоглобина О2 имеет сигмоидальную форму. Т.о., способность гемоглобина связывать О2 зависит от того, содержатся ли в данном тетрамере другие молекулы О2. Если да, то последующие молекулы О2 присоединяются легче. Для гемоглобина характерна кинетика кооперативного связывания, благодаря которой он связывает максимальное количество О2 в легких и отдает максимальное количество О2 при тех значениях Р О2 , которые имеют место в периферических тканях.
Сродство гемоглобинов к О2 характеризуется величиной Р50- значением парциального давления О2, при котором наблюдается полунасыщение гемоглобина кислородом. Например, для HbA Р50 = 26 мм рт.ст., а для HbF - 20 мм рт. ст. Благодаря этой разнице гемоглобин F отбирает кислород у HbA, находящегося в плацентарной крови.
2) У олигомерных белков имеется специальный центр, отличный от активного, к которому присоединяется аллостерический лиганд. Этот центр получил название аллостерического.
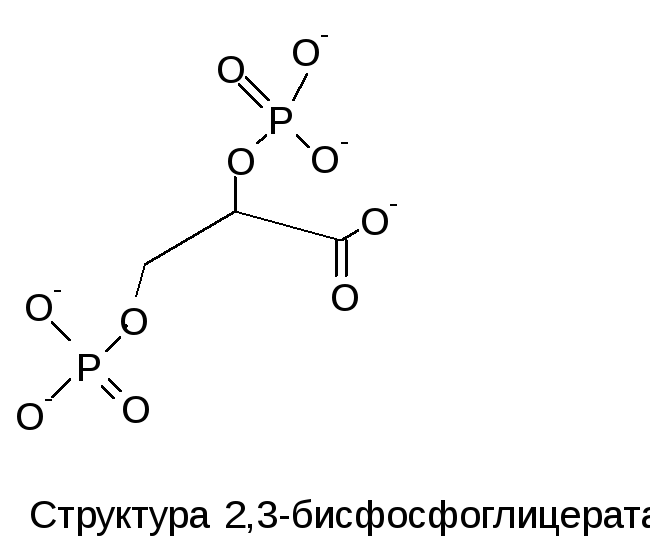
Рассмотрим аллостерическую регуляцию на примере HbА. В центре молекулы HbА находится полость, которая является местом присоединения 2,3-ДФГ. Это соединение накапливается в эритроцитах при недостатке О2. Непосредственным предшественником служит 1,3-дифосфоглицерат - промежуточный продукт гликолиза. 2,3-ДФГ имеет сильный отрицательный заряд и взаимодействует с 5 положительно заряженными группами аллостерического центра. В результате взаимодействия образуется 5 дополнительных ионных связей, что снижает сродство Hb к кислороду и облегчает его отдачу в ткани.
Кооперативные изменения конформации олигомерных белков составляют основу механизма регуляции функциональной активности не только Hb, но и большого числа других белков, в том числе аллостерических ферментов.
5. Изофункциональные белки
Белок, выполняющий определенную функцию в клетке, может быть представлен несколькими формами - изофункциональными белками или изобелками. Такие белки хоть и выполняют одинаковую функцию, но отличаются константой связывания, что приводит к некоторым различиям в функциональном отношении. Например, в эритроцитах человека обнаружено несколько форм гемоглобина: HbA (96%), HbF (2%), HbA2 (2%). Все гемоглобины представляют собой тетрамеры, построенные из протомеров , , , (HbA -22, HbF - 22, HbA2 - 22). Все протомеры сходны между собой по первичной, вторичной и третичной структурам. Все формы гемоглобинов предназначены для переноса кислорода в клетки тканей, но HbF, например, имеет большее сродство к кислороду, чем HbA. HbF характерен для эмбриональной стадии развития человека. Он способен отнимать кислород у HbA, что обеспечивает нормальное снабжение кислородом плода.
Изобелки - это результат наличия более чем одного структурного гена в генофонде вида.