

4.2.3. Способность к специфическим взаимодействиям как основа биологической активности белков
В основе функционирования белка лежит его способность к взаимодействию с каким-либо другим веществом - лигандом. Лигандом может быть как низкомолекулярное вещество, так и макромолекула, в том числе и другой белок. Лиганд присоединяется к определенному участку глобулярной молекулы - активному центру. Активный центр формируется в ходе образования третичной структуры белка, поэтому при денатурации белка (когда разрушается его третичная структура) белки теряют свою активность. Специфичность взаимодействия белка и лиганда объясняется комплиментарностью пространственной структуры активного центра и молекулы лиганда. Взаимодействие между белком P и лигандом L описывается уравнениями:
P + L PL
![]()
Ксв определяет сродство белка к данному лиганду, чем больше Ксв, тем больше сродство.
На специфичности взаимодействия белков и лигандов основан метод разделения белков - аффинная хроматография.
4.2.4. Ингибиторы функций белков
Учитывая, что различные лиганды отличаются Ксв, всегда можно подобрать вещество, похожее по структуре на природный лиганд, но имеющий большее значение Ксв с данным белком. Например, СО имеет Ксв в 200 раз больше, чем О2 с гемоглобином, поэтому достаточно 0,1% СО в воздухе, чтобы заблокировать большое количество молекул гемоглобина. По такому же принципу действуют многие лекарства. Например, дитилин.
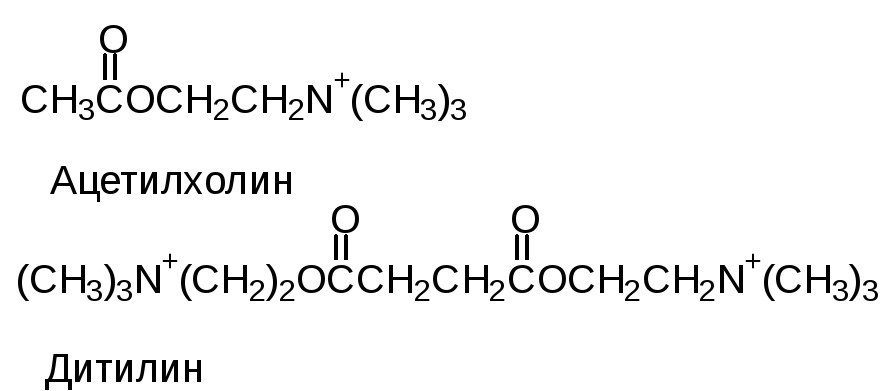
Ацетилхолин - медиатор передачи нервных импульсов на мышцу. Дитилин, имея большую Ксв, блокирует рецептор, с которым связывается ацетилхолин, и вызывает эффект парализации.
4.2.5. Четвертичная структура белка.
Под четвертичной структурой белка подразумевают способ укладки в пространстве отдельных полипептидных цепей, обладающих первичной, вторичной и третичной структурой и формирование единого в структурном и функциональном отношении макромолекулярного образования. Каждая отдельная полипептидная цепь (протомер или субъединица) не обладают биологической активностью, а образовавшаяся молекула - олигомер обладает биологической активностью. Четвертичная структура белка уникальна, как и другие уровни организации. Четвертичная структура поддерживается нековалентными взаимодействиями между контактными площадками протомеров.
Олигомерные белки обладают особыми свойствами, которых нет у белков, не имеющих четвертичной структуры:
1) наличием кооперативного взаимодействия между субъединицами,
2) наличием аллостерической регуляции
Четвертичная структура белков - еще один пример удивительной мудрости природы. Докажем это на примере функционирования двух белков: Mb – белка мышц и Hb – белка эритроцитов. Mb и протомеры Hb имеют сходную первичную, вторичную и третичную структуры. Оба белка являются гемопротеинами, обладающими способностью связывать кислород. При этом миоглобин обладает только третичной структурой и гемоглобин (Hb) является тетрамером и содержит по две цепи двух разных видов: и . Их длина примерно одинакова (- 141 а.к., - 146 а.к.).
Простетической группой этих белков является гем, который расположен в гидрофобном кармане пептидной цепи каждого протомера, т.е. окружен неполярными остатками, за исключением 2-х остатков гистидина, расположенных по обе стороны плоскости гема. С одним из них (проксимальным гистидином) Fe2+ связано координационно по 5 координационному положению. Второй (дистальный) гистидин расположен почти напротив проксимального, но несколько дальше, поэтому 6-ое координационное положение Fe2+ остается свободным. В неоксигенированном миоглобине или протомере гемоглобина атом железа выступает из плоскости кольца в направлении проксимального гистидина на 0,03 нм. В оксигенированном миоглобине (протомере гемоглобина) кислород занимает 6-ое координационное положение атома железа, при этом Fe2+ не меняет степени окисления. Такого эффекта позволяет достичь гидрофобное белковое окружение гема, которое не позволяет кислороду слишком сблизиться с железом, чтобы его окислить. Железо, координируя О2, выступает из плоскости кольца лишь на 0,01 нм. Т.о., оксигенирование миоглобина сопровождается смещением атома железа и, следовательно, проксимального гистидина и ковалентно связанных с ним аминокислотных остатков в направлении плоскости кольца. В результате белковая глобула меняет конформацию. Эти изменения в гемоглобине сопровождаются разрывом связей между протомерами, что облегчает связывание следующих молекул О2. В результате сродство Hb к четвертой молекуле О2 в 300 раз больше, чем к первой. Это явление называется кооперативностью изменения конформации протомеров.