

3 курс / Топографическая анатомия и оперативная хирургия / Хирургическая_анатомия_венечных_артерий,_Бокерия_Л_А_,_Беришвили
.pdf2.Эмбриологические предпосылки ветвления
иследования венечных артерий
Прежде чем оценить характер следования основных стволов эпикардиальных ВА, представляется логичным определиться терминологически, поскольку в литературе мнения на этот счет весьма противоречивы. Для упорядочения наших представлений о том, что из себя представляет ВА в зрелом сердце, следует проследить их эмбриогенез.
Забегая вперед, было бы уместным отметить, что следование ВА в сердцах в норме и при ВПС изменяется мало. Основные изменения в строении венечно-артериального дерева заключаются в различиях в отхождении и ветвлении ВА. Поэтому сначала ко ротко остановимся на ветвлении ВА.
Согласно данным, приведенным выше, ВА закладывается в виде колец в области межсегментных соединений (см. рис. 1). После петлеобразования сердца кольца ВА сходятся в области бульбовентрикулярной складки (см. рис. 2). Их свободные концы веерообразно расходятся, охватывая примитивный желудочек и бульбус (луковицу). Кольца А охватывают бульбус, а кольца С и Е - желудочек. Кольцо В соединяется с ними в двух точках (Вх - слева и By - справа) в области слияния вокругбульбарных
(А) и вокругжелудочковых колец (см. рис. 3). Венечно-артериальное сплетение изоли ровано от системной циркуляции. Согласно нашим данным, при атрезии правого атриовентрикулярного отверстия синусная часть, равно как и задняя межжелудочковая перегородка, не сформирована. На фрагменте Б рисунка 3 воспроизведен срез сердца с атрезией правого атриовентрикулярного отверстия. Как следует из рисунка, сердце при данном пороке мало чем отличается от сердец с примитивной бульбовентрикуляр ной петлей. Примитивный желудочек сохранен и полностью принимает весь приток из предсердий. Однако общий артериальный ствол уже разделен на аорту и легочную артерию. Как и в нормальном сердце, от 2-го лицевого синуса аорты отходит левая ВА, делящаяся на ПМЖВ, следующую в передней межжелудочковой борозде (см. рис. 9, фрагмент А, круг В), и ОВ, располагающуюся в левой атриовентрикулярной борозде (рис. 3,Б - круг С, рис. 9,А - круг С).
Как уже отмечалось выше, при достаточном развитии желудочковой камеры вокругжелудочковые круги D, Е (см. рис. 3,Б) разрываются. Аналогично, с ростом под легочного инфундибулума разрываются вокругбульбарные круги (рис. 3,Б - круг А). Поэтому правая ВА, отходящая от 1-го лицевого синуса аорты, может соединиться с левой только посредством круга В (см. рис. 9). Однако в силу роста выводной камеры точка соединения круга В с кругом С (см. рис. 3,Б - точка 3) уже не совпадает с точкой соединения кругов А и С (см. рис. 3,Б - точка 2).
Левая ветвь круга В, отходящая от точки 1 (см. рис. 3,Б), следующая вдоль перед ней межжелудочковой борозды, представляет собой ПМЖВ. Правая ветвь круга Вр отходящая от точки 3, представляет собой ветвь острого края. ЗМЖВ в сердцах с атре зией правого атриовентрикулярного отверстия не сформирована. В этих сердцах от сутствуют задняя перегородка и синусная часть правого желудочка (И. И. Беришвили и др., 1982) и, соответственно, нет задней межжелудочковой борозды. Более того, оба предсердия при данном пороке дренируют кровь в левый желудочек. Крест сердца, об разованный межжелудочковой и межпредсердной перегородками и атриовентрику лярной бороздой, отсутствует. Ветвь острого края отходит в проекции, несоответству ющей даже условному кресту сердца. Поэтому, несмотря на то, что S. Bharati и соавт. (1977) полагают, что эта ветвь представляет собой ЗМЖВ, мы разделяем мнение
R.Anderson и A. Becker (1981) и не можем определить ее как ЗМЖВ.
Всердцах с единственным желудочком (см. рис. 13, В) левожелудочкового типа же лудочковая камера сформирована полностью, причем включает в себя и синусную часть правого желудочка. Правый желудочек при этом пороке не сформирован и представлен только его выводным отделом (бульбусом). Как правило, отсутствует задняя перегород ка, хотя в редких случаях она может формироваться, но быть гипопластичной. И в этом
76

случае левая ВА отходит от 2-го лицевого синуса и в точке 1 (см. рис. 13,В) делится на ПМЖВ и ОВ. ПМЖВ представляет собой левую ветвь круга В (Вх - рис. 13,В) и следует вдоль передней межжелудочковой борозды (рис. 29, 30). Вследствие достаточного роста желудочковой камеры круги D и Е, как правило, разрываются (рис. 31), но иногда со хранены (рис. 32). Понятно, что при отсутствии задней перегородки ЗМЖВ в этих серд цах чаще всего отсутствует. Ветвь острого края (рис. 33) представляет собой правую часть фетального круга В, и она чаще всего разорвана. Сохранность этого круга, разде ляющего бульбарную и желудочковую камеры, равно как и кругов Вьессена (см. рис. 13,В), обусловлена развитием выпускника. При достаточном развитии камеры кру ги Вьессена разрываются, хотя могут быть представлены его остатками (см. рис. 29); при выраженном же недоразвитии - практически полностью сохраняются. При этом может сохраниться и сообщение между правой и левой сторонами (ВОК и ПМЖВ) круга В.
Рис. 29. Венечные артерии в сердце с единст венным желудочком.
Препарат сердца. Вид спереди справа.
Сохранение остатков круга Вьессена (ярусная конусная артерия - КА, правожелудочковые ветви - ПЖВ) перед ней межжелудочковой ветви (ПМЖВ).
А - аорта.
Рис. 30. Венечные артерии в сердце с единственным желудочком.
Препарат сердца. Вид спереди слева. Нормаль ное развитие ветвей левой венечной артерии: ПМЖВ, ОВ, ДВ, ВТК.
ПМЖВ - передняя межжелудочковая ветвь, ОВ - огибающая ветвь, ДВ - диагональная ветвь, ВТК - ветвь тупого края, А - аорта.
Как уже отмечалось (И. И. Беришвили и др., 1991), в основе формирования правого желудочка лежит формирование и развитие сначала его синусной части, естественно, с одновременным развитием и его бульбарной части. Завершается формирование право го желудочка образованием задней межжелудочковой перегородки. В результате таких особенностей формирования правожелудочковой камеры бульбовентрикулярное отвер стие, ранее разделявшее бульбус и примитивный желудочек, в сформированном ПЖ разделяет приточный и выводной отделы. В соответствии с этими изменениями меня ют свое расположение и кольца первичного венечно-артериального сплетения. В силу адекватного развития подлегочного инфундибулума круг Вьессена разрывается, и ко нусная артерия становится ветвью правой ВА (см. рис. 9,А), питающей область па риетального внедрения над желудочкового гребня. Ветвь острого края (правая часть круга В) смещается кзади, к острому краю сердца (к границе между диафрагмальной
77

Рис. 31. Венечные артерии в сердце с единственным же лудочком.
Препарат сердца. Вид сзади на же лудочковую камеру.
Разрыв вокругжелудочковых кру гов D и Е (показано стрелками).
Рис. 33. Венечные артерии в сердце с единственным желудочком.
Препарат сердца. Вид сбоку справа.
То, что в нормальном сердце является ЗМЖВ, в данном сердце представляет собой ветвь острого края (ВОК). ПВА - правая венечная артерия.
Объяснение в тексте.
Рис. 32. Венечные артерии в сердце с единственным желудочком.
Препарат сердца. Вид сзади на желудочковую камеру.
Сохранение вокругжелудочковых кругов D и Е (показано стрелками).
и передней стенками правого желудочка) и, от ходя от правой ВА уже в регионе притока, (рис. 9,А и 33), теряет всякую связь с КА.
Продолжающаяся далее к кресту сердца правая ВА (чаще всего круг Е) (см. рис. 9,Б) в области проекции задней межжелудочковой перегородки формирует ЗМЖВ. Эта ветвь рас полагается в задней межжелудочковой перего родке и следует перпендикулярно книзу от атриовентрикулярной борозды строго вниз от креста сердца (см. рис. 13,Г - точка 4).
Таким образом, во всех без исключения сердцах ПМЖВ сформирована и располагает ся в передней межжелудочковой борозде.
Ее отсутствие - аномалия.
Говорить о наличии ЗМЖВ следует только в сердцах со сформированными желудочками, со держащих заднюю межжелудочковую перего родку. Атрезия правого атриовентрикулярного отверстия и сердца с единственным желудочком (левожелудочкового типа) ЗМЖВ не имеют.
В нормальных сердцах ЗМЖВ располагает ся в задней межжелудочковой перегородке, сле дуя перпендикулярно вниз от креста сердца.
Отсутствие ЗМЖВ делает безопасным накладывание швов при септации сердец с единственным желудочком, но, к сожале-
78
нию (ввиду кровоснабжения бульбовентрикулярной складки ветвью острого края правой ВА), не позволяет рассчитывать на пластическое расширение рудиментар ной камеры при атрезии правого атриовентрикулярного отверстия (создание при точного отдела и «правого желудочка» за счет заплаты или использования метода кардиомиопластики).
Близость ВОК и ПМЖВ по ангиограммам свидетельствует о выраженной гипопла зии подлегочного инфундибулума. При этом сохраняется круг Вьессена. Сохранение круга Вьессена возможно и при нормально развитом инфундибулуме. Сохранение кру га Вьессена при нормально развитом подлегочном инфундибулуме - свидетельство не достаточности кровотока в системе левой ВА (аномальное отхождение левой ВА от ЛА, окклюзия или стеноз проксимального отдела ПМЖВ или ствола ЛА).
Как следует из приведенных данных, эмбриологически круг Вьессена -единствен ный первоначально закладываемый путь соединения систем правой и левой ВА. По этому его функционирование - свидетельство неэффективности кровотока по системе левой ВА. Учитывая отсутствие эффективной анатомической связи между ветвями ле вой ВА, при операциях реваскуляризации миокарда нельзя рассчитывать на достаточ ный кровоток через круг Вьессена. Этот путь коммуникаций эффективен только для кровоснабжения ПМЖВ. Об этом, в первую очередь, следует думать при аномальном отхождении левой ВА от ЛА. В этих случаях всегда необходимо планирование опера ции, позволяющей достичь адекватного кровотока по ОВ левой ВА. Из сказанного так же ясно, что реваскуляризация ветвей ОВ - важнейший аспект проблемы лечения больных ишемической болезнью сердца.
3.Следование правой и левой венечных артерий
иих основных эпикардиальных стволов
Как показало сопоставление материала по изучению следования правой и левой ВА и их основных эпикардиальных стволов с различными факторами, определяющи ми посегментное строение сердца (сторона петлеобразования сердца, атриовентрикулярная конкордантность или дискордантность, сторона ротации конотрункуса), при двухкоронарной системе кровоснабжения сердца огибающие ветви и правой и левой ВА всегда следуют в атриовентрикулярной борозде, принадлежащей одноименному (правому или левому) желудочку, и это - закономерность.
В сердцах с единственной ВА следование эквивалентов правой и левой ВА тоже подчинено указанной закономерности.
Приведенная закономерность в полной мере относится и к сердцам с гипоплазией желудочков, а применительно к левой ВА - и к сердцам с атрезией правого атриовен трикулярного отверстия и единственному желудочку левожелудочкового типа.
Несмотря на то что сегодня уже понятно, что при большинстве ВПС без наруше
ний в строении конуса ветвление ВА не отличается от такового в норме |
(И. И. Бе- |
|||
ришвили и М. Н. Вахромеева, 1990; |
Н. Neufeld |
and A. Schneeweiss, 1983; |
Th. Lesser |
|
and A. Schneider, 1990), установлено, |
что при |
пороках конотрункуса |
изменяется |
|
привычное ветвление ВА (R. Dabizzi et al., 1980; A. Gittenberger-de Groot |
et |
al., 1983; |
||
J. Berry et al., 1988). Кроме того, доказано, что несмотря на то что нисходящие эпикардиальные стволы ВА меняют источник питания, анатомия дистальных ВА при них не изменена, и кровоснабжение миокарда не нарушается (В. Berry and D.McGoon, 1973; J.Meyer et al., 1975; M. De La Cruz et al., 1989, 1990). Объяснения механизмов таких перестановок в литературе не даны и, в принципе, из данных ли тературы непонятны.
Как показал анализ наших данных (см. рис. 21), различия в строении ВА в норме и при пороках конотрункуса заключаются только в отхождении крупных венечно-ар- териальных стволов от правой или левой ВА (изменение типа ветвления - например,
75»

отхождение ПМЖВ от ПВА), либо отхождении самих ВА (правой или левой) от лице вых синусов аорты (например, отхождение правой ВА от 2-го лицевого синуса аорты, а левой - от 1-го). В чем же причина таких изменений?
Согласно нашей концепции формирования венечно-артериальной системы, нисхо дящие ВА, представляющие сохранившиеся эквиваленты веерообразно расходящих ся сегментов колец первичного венечно-артериального сплетения, являются комму никантами между перитрункальным кольцом и миокардом. Сформировавшись и со хранившись во всех сердцах со сформированными желудочками, они обеспечивают одинаковую (и адекватную) перфузию миокарда во всех сердцах (и в норме и при ВПС). Другое дело, что они не во всех сердцах питаются из идентичных источников.
Смена источника питания обусловлена использованием различных фрагментов перитрункального кольца, что определяется главным образом развитием инфундибулума переднего артериального сосуда и разворотом оси конотрункуса.
Отсюда - различия в строении венечно-артериальной системы в сердцах с нор мальным разворотом конотрункуса и ненормальным. Но отсюда же и нормальность перфузии миокарда во всех сердцах.
Таким образом, в правой атриовентрикулярной борозде всегда следует правая ВА, в левой - ОВ левой ВА. Отсутствие правой ВА в литературе не описано, отсутствие ОВ - редчайшая аномалия (Г. И. Цукерман и др., 1976; R. Bestetti et al., 1985).
Как известно, доминантность кровоснабжения сердца определяется тем, какая и:: ВА кровоснабжает ЗМЖВ.
Доминантность ВА нами изучена в 300 сердцах с разворотом конотрункуса, харак терным для нормальных сердец. Согласно нашим данным (табл. 1), в 75% случаев до минантна правая ВА, в 2 1 % - левая и только в 4% случаев определяется сбалансиро ванный тип кровоснабжения сердца. Эти данные противоречат как материалам, при веденным в отечественной литературе (С. В. Самойлова, 1970; В. В. Кованое и Т. Н.
Аникина, 1974), |
так |
и |
данным иностранных авторов (R.Anderson and A. Becker, 1981; |
К. Venkataraman |
et |
al., |
1988). |
Т а б л и ц а 1
Распределение типов доминантного кровоснабжения в сердцах с «нормальным» разворотом конотрункуса
Как свидетельствуют данные, представленные в табл. 1, чем меньше разворот конотрункуса по часовой стрелке, тем больше вероятность существования левого типа доминантности кровоснабжения сердца, и - наоборот. Эти данные позволяют заподо зрить, что формирование того или иного типа доминантности кровоснабжения сердца в определенной степени связано с процессом вращения конотрункуса и определяется степенью его разворота.
80
Здесь, по-видимому, следует подчеркнуть, что в подавляющем большинстве случа ев доминантность никак не определяет перфузию миокарда, поскольку независимо от типа доминантности все участки миокарда кровоснабжаются адекватно. Доминант ность кровоснабжения миокарда приобретает особое значение лишь при патологии ве- нечно-артериальной системы (отхождении левой ВА от ЛА или окклюзии ствола ле вой ВА, например), поскольку в этих случаях именно доминантностью определяется инфарцирование задней стенки левого желудочка; ею же определяется и то, будет ли страдать зона кровоснабжения проводящей системы сердца.
Как уже отмечалось выше, ПМЖВ сформирована во всех сердцах и следует в пе редней межжелудочковой борозде. Отсутствие ПМЖВ - аномалия.
При отсутствии задней перегородки говорить о доминантности кровоснабжения сердец с атрезией правого атриовентрикулярного отверстия или единственным желу дочком представляется неправомочным. По-видимому, этот термин следует использо вать только в сердцах со сформированными желудочками (и в норме и при ВПС).
Поскольку практически во всех этих сердцах в атриовентрикулярных и межжелу дочковых бороздах следуют крупные артериальные стволы, перфузия миокарда не страдает ни в норме, ни при ВПС. В этом - идентичность кровоснабжения сердца в нор ме и при ВПС. Это обусловлено идентичностью закладки ВА (в атриовентрикулярных и межжелудочковых бороздах) во всех сердцах со сформированными желудочками.
Таким образом, существуют следующие закономерности следования ВА.
1.Сторона следования ОВ правой или левой ВА определяется стороной расположе ния одноименного желудочка. Она всегда располагается в атриовентрикулярной борозде, образованной этим желудочком.
2.Нисходящие ветви ВА располагаются в одноименных межжелудочковых бо роздах:
а) ПМЖВ располагается в передней межжелудочковой борозде; б) ЗМЖВ формируется только в сердцах со сформированными желудочками и
располагается в задней межжелудочковой борозде (и тем самым, как прави ло, свидетельствует о наличии сформированных желудочков в сердце).
4. Ветвление венечных артерий
Проанализировав различные варианты отхождения и ветвления ВА в 512 сердцах, мы пришли к однозначному выводу, что в принципе все существующие варианты от хождения и соединения (ветвления) ВА можно сгруппировать в несколько групп. В од ну группу могут попасть и нормальные сердца, и сердца с ВПС, причем разными.
Сравнив указанные варианты в 500 сердцах с нормальным отхождением ВА от аортальных синусов и обнаружив, что в принципе группы различаются только по отхождению от синусов аорты либо набору сегментов, участвующих в образовании ве- нечно-артериальной системы сосудов, мы пришли к выводу, что имеется некая стан дартная исходная организация венечно-артериального сплетения (рис. 34), которая в процессе окончательного формирования сердца использует то одни, то другие сегмен ты общей исходной системы, причем в одной группе - обязательно одни и те же.
Таким образом, согласно нашим данным, первичная закладка венечно-артериаль- ной системы константна. Она идентична во всех сердцах.
В дальнейшем, в процессе формирования окончательной (зрелой) системы ВА, часть сегментов первичного венечно-артериального сплетения сохраняется, а другая часть - редуцируется.
Возможность формирования различных формул соединения сегментов ВА в прин ципе, как оказалось, в пределах одной стороны петлеобразования сердца ограничена 11 вариантами.
При первом типе соединения ВА правая ВА отходит от 1-го лицевого синуса аорты, а левая - от 2-го лицевого синуса, делясь на ПМЖВ и ОВ.
S1
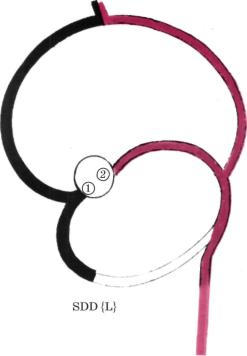
|
В этой группе конусная и адвентициальная |
|
|
артерии, равно как и артерия острого края, мо |
|
|
гут отходить от самостоятельного 1-го лицевого |
|
|
синуса аорты. От 2-го лицевого синуса аорты са |
|
|
мостоятельно могут отходить и ветви левой ВА |
|
|
(ОВ и ПМЖВ). |
|
|
Второй тип соединения ВА отличается от пер |
|
|
вого наличием крупной КА, больше характерного |
|
|
для сердец с относительным недоразвитием под |
|
|
легочного инфундибулума. |
|
|
Третий тип соединения ВА характеризуется |
|
|
сохранением передней части круга Вьессена и в |
|
|
нашей серии встретился только в сердцах с тетра |
|
|
дой Фалло и ОАЛА ПЖ со стенозом легочной ар |
|
|
терии, то есть в случаях с выраженной гипопла |
|
|
зией подлегочного инфундибулума. При этом ти |
|
|
пе соединения ВА ПМЖВ кровоснабжается из си |
|
|
стем обеих (правой и левой) ВА и пересекает вы |
|
|
водной отдел ПЖ . Правая ВА, отходящая от 1-го |
|
|
лицевого синуса аорты, отдает КА, соединяющу |
|
|
юся с ПМЖВ. Левая ВА отходит от 2-го лицевого |
|
Рис. 34. Схема первичного венечно- |
синуса аорты и делится на ОВ и ПМЖВ. Первая |
|
огибает митральный клапан и образует ветвь ту |
||
артериального сплетения в сердцах с |
||
бульбовентрикулярной петлей (с сег |
пого края. Вторая же соединяется с К А и далее |
|
ментарной формулой SDD [L]). |
следует в передней межжелудочковой борозде. |
|
Здесь, а также на рисунках 35 и 108 показано, |
Четвертый тип соединения ВА встречается |
|
что из темного фрагмента сплетения заклады |
чаще, чем третий, и характеризует выраженную |
|
вается система правой ВА, отходящей в нор |
||
гипоплазию подлегочного инфундибулума и, как |
||
мальных сердцах от 1-го (1) лицевого синуса |
||
аорты, из красного — система левой ВА, отходя |
правило, легочного кольца. ПМЖВ в этих случа |
|
щей в нормальных сердцах от 2-го (2) лицевого |
ях является ветвью правой ВА и пересекает вы |
|
синуса аорты. Светлым фрагментом показана |
||
водной отдел ПЖ . Она не соединяется с системой |
||
передняя часть круга Вьессена, формирующая |
||
конусную артерию, правожелудочковую ветвь |
левой ВА, представленной только ОВ и ее ветвя |
|
передней межжелудочковой артерии и некото |
ми. ОВ отходит от 2-го лицевого синуса аорты. |
|
рые редкие варианты ВА. |
||
Пятый тип соединения ВА имеет единствен |
||
|
||
|
ную ВА, отходящую от 2-го лицевого синуса аор |
ты, а в сердцах с нерассосавшейся бульбовентрикулярной складкой - самостоятельное отхождение ПМЖП от 1-го лицевого синуса аорты. ПМЖВ в этих случаях не соедине на с системой левой ВА.
Не вдаваясь в детальное описание всех типов соединения ВА, представленных на рис. 21, считаем уместным подчеркнуть однотипность изменений в соединении эле ментов венечно-артериальной системы. По мере увеличения разворота оси конотрун куса (по или против часовой стрелки) эти изменения носят строго определенный ха рактер, заключающийся в равноизменении при равноудаленности.
Анализ этих данных показал, что в принципе существует некоторая исходная по зиция конотрункуса, при вращении от которой по или против часовой стрелки образу ется шесть абсолютно симметричных пар соединения ВА. Причем при вращении коно трункуса по часовой стрелке последняя пара образована нормальным сердцем, а про тив часовой стрелки - АКМАЛА.
Представляется важным отметить еще одну закономерность, характеризующую сердца с небольшим разворотом конотрункуса от исходной позиции. Здесь формируется специфический класс соединения ВА с единственной ВА. Иными словами, 8 из 11 фор мул соединения ВА обеспечивают формирование двухкоронарной системы кровоснаб жения сердца, а 3 - однокоронарной. Причем варианты однокоронарного типа кровооб ращения сердца занимают центральное положение, а двухкоронарного - краевое.
82

Рис. 35. Схема симметричности закладки первичного венечно-артериального сплетения в серд цах с правой (сегментарная формула SDD [L]) и левой (сегментарная формула SLL [D]) бульбовентрикулярной петлей.
Представляют интерес и материалы по сопоставлению типов соединения ВА в сердцах с различной стороной петлеобразования сердца. Оказалось, что при равной степени ротации конотрункуса тип соединения ВА при различных петлях также абсо лютно симметричен (рис. 35). Отличия между ними заключаются только в изменении формирования петли справа или слева. Никакие прочие факторы не определяют стро ения венечно-артериальной системы.
Таким образом, первичная закладка системы венечно-артериальных сосудов идентична во всех сердцах и не зависит ни от типа симметрии предсердий и органов брюшной полости, ни от потенциального конечного типа межартериальных взаимо
отношений, |
ни от нормальности либо порочности окончательного формирования |
||
сердца. |
Она |
определяется |
только стороной образования булъбовентрикулярной пет |
ли и в |
сердцах с L-петлей |
полностью обратна (зеркальна) таковой при D-петле. |
|
Приведенные материалы свидетельствуют о наличии симметрии в строении ВА. Причем симметричны и равноудалены от исходной позиции по мере ротации коно трункуса формулы соединения ВА и симметричны одинаковоудаленные от фронталь ной (нулевой) линии формулы при разных петлях. Понимание этих моментов важно с практической точки зрения.
Так, понимание симметрии в пределах одной петли, например, позволяет знать, что первые две формулы имеют такое расположение ВА, при котором рассечение вы водного отдела ПЖ не лимитировано, а последующие четыре формулы не позволяют сделать это, так как выводной отдел ПЖ пересекается крупной В А.
Первичная венечно-артериальная система складывается во всех сердцах одинако во. Она образуется и полностью сформирована уже при фетальной жизни.
Дальнейшие ее изменения определяются процессом формирования сердца. Важ нейшим из факторов, определяющих формирование различных вариантов соединения (ветвления) венечно-артериальной системы, является ротация конотрункуса. Причем тут важно понять, что в процессе формирования венечно-артериальной системы ВА не меняют своего положения. В процессе ротации конотрункуса к лицевым синусам аор ты прилегают различные отделы перитрункального кольца, что и определяет вариан ты отхождения ВА. Более того, при вращении конотрункуса формирующаяся система ВА использует различные сегменты перитрункального кольца. Это последнее - веду щий фактор, определяющий варианты ветвления ВА.
Отсюда понятно, что тип соединения ВА сегментов первичной венечно-артериаль ной системы в каждом сердце детерминирован. Он обусловлен сектором разворота
83
оси конотрункуса. Варианты измененного соединения ВА для каждого сектора оста новки ротации конотрункуса (при отсутствии прочих нарушений, например, в стро ении перегородок сердца) идентичны во всех сердцах и поэтому закономерны.
Завершая разговор об образовании той или иной формулы соединения ВА, хо чется отметить, что складывание определенной системы ВА обусловлено парал лельными изменениями в пространственной ориентации множества структур. Так, например, одно только изменение геометрии перегородок сердца может привести к аномальному отхождению ВА от легочной артерии, а отсутствие выравненности комиссур - к формированию аномальной единственной ВА, то есть формированию единственной ВА в том секторе, где она не должна быть. Но тут важно знать, что возможность формирования различных формул соединения сегментов ВА чаще все го ограничена 11 вариантами (при согласованном формировании всех структур, от вечающих за конечное формирование венечно-артериального сплетения в пределах данного сектора), вероятность же формирования аномалий крайне низка, но воз можных вариантов много.
Таким образом:
A. Сохранение или облитерация отдельных сегментов венечно-артериальной сис темы и конкретный тип их соединения в зрелом (нормальном или прочно сформиро ванном) сердце обусловлены главным образом направлением и процессом ротации конотрункуса (по часовой стрелке или против нее).
Б. В зависимости от степени ротации конотрункуса принципиально для каждой петли (D и L) существует 11 возможных вариантов (формул) соединения фрагментов венечно-артериальной системы.
B. Каждый тип соединения (формула) сегментов венечно-артериальной системы обусловлен и характеризуется строго определенным взаимоотношением аорты и ле гочной артерии на уровне полулунных клапанов (то есть имеет свои границы секто ра вращения конотрункуса) и не зависит от нормальности или порочности формиро вания данного сердца.
Г. Каждый тип соединения венечно-артериальной системы имеет свой вариант нормальности (закономерность) соединения стволов ВА, и нарушение этой системы для каждой отдельной формулы является аномалией.
Д. При первых четырех и последних четырех из 11 типов соединения (формул) сегментов венечно-артериальной системы в норме складывается двухкоронарная система кровообращения, а при промежуточных трех - однокоронарная.
Е. Для отдельных типов соединения (формул) ВА, как и для определенного сек тора пространственных взаимоотношений аорты и легочной артерии на уровне полу лунных клапанов, то есть при отсутствии ротации конотрункуса относительно при митивного желудочка, имеющего место на ранних этапах формирования сердца, на личие единственной ВА является не только нормальным, но и обязательным. В этом секторе при отсутствии сопутствующих нарушений формирование двухкоронарнои системы кровообращения сердца невозможно.
Ж. Аномалии ВА чаще всего обусловлены либо самостоятельной дополнительной ротацией аорты и легочной артерии в пределах данного сектора их пространствен ных взаимоотношений и, соответственно, несовпадением лицевых полулунных за слонок и комиссур, либо особенностями перегородок сердца.
5. Окончание венечных артерий
Не располагая достаточным количеством секционных наблюдений вариантов и аномалий окончания ВА, мы можем ограничиться констатацией только одной законо мерности: в норме все основные эпикард иальные стволы венечных артерий должны заканчиваться интрамиокардиально, участвуя в формировании капиллярного русла.
84
6. Обсуждение
Высказанные в работе соображения по части обсуждаемых вопросов в литературе имеют определенное подтверждение. Правда, эти данные большей частью касаются изучения коллатерального кровообращения и, понятно, не увязаны с общей концеп цией строения всей венечно-артериальной системы.
Так, например, известно, что:
1. Венечно-артериальная циркуляция образуется и полностью сформирована уже
при фетальной |
жизни |
(W.Laurie |
and |
J.Woods, 1958; L.Reiner et al., 1961; |
A. Rather et al, |
1973; M. |
Miyzaki and |
M. |
Kato, 1988). |
2. Между ветвями обеих (правой и левой) систем ВА существует врожденная сеть
коммуникаций (M.Vastesaeger et al., |
1957; M.Zanchi and L.Locatelli, 1958; |
T. James, 1970; D. Gregg and R. Patterson, |
1980; D. Wahr et al., 1985). |
3.С возрастом, при отсутствии необходимости в ее функционировании, часть пер вичной фетальной венечно-артериальной системы и коммуникации между ее отдельными частями претерпевают инволюционные изменения (L. Reiner et al., 1961).
4.Изменения эти носят обратимый характер, позволяя превращаться из нефункционирующей в функционирующую сеть сообщений ВА при возникновении не
обходимости ее существования (D.Levin, 1974; L.Friedman et al., 1982; J.Hansen, 1989).
5.При некоторых ВПС инволюции межкоронарных анастомозов не происходит: их сохранение обусловлено изменениями в строении сердца, магистральных со
судов и ВА (R. Von Wuttrich, 1951; S. Van Praagh et al., 1982).
Наши рассуждения об общей концепции строения венечно-артериальной системы не расходятся с этими данными. Но в них, понятно, акцент поставлен на особенностях и закономерностях всей системы, а не на путях формирования межкоронарных анас томозов. Хотя и формирование анастомозов является фрагментом образования и раз вития всей венечно-артериальной системы, и в этом смысле мы, естественно, опира лись на материалы, представленные в литературе.
Но ставя перед собой цель выявить закономерность строения венечно-артериаль ной системы, мы не могли ограничиться описанием особенностей анатомии ВА в нор мальном сердце либо описанием аномалий В А при ВПС. Не могли мы ограничиться и простой констатацией частоты аномалий ВА при ВПС. Поэтому мы отказались от ана лиза аномалий и оценки вариантов ВА в общепринятом смысле. Для отхода от стерео типных подходов в изучении ВА мы попытались проанализировать зависимость стро ения ВА от целого ряда факторов. Поскольку уже на этапе предварительного анализа данных была обнаружена связь между строением венечно-артериальной системы и ро тационными особенностями конотрункуса, мы поставили перед собой цель изучить возможно более широкий спектр сердец в пределах всего диапазона ротации коно трункуса, причем в пределах обеих сторон петлеобразования сердца. Такой подход, как оказалось, предоставляет исследователю широкие возможности для многочислен ных сопоставлений и обобщений.
Для выяснения всех основных причин, определяющих формирование венечно-ар териальной системы, мы, кроме того, изучали: 1) характер межартериальных взаимо отношений; 2) тип желудочково-артериальной связи; 3) отношение магистральных артерий к основанию сердца и его перегородкам; 4) тип межжелудочковых взаимоот ношений; 5) сегментарную формулу сердца.
Первоначальный анализ данных показал существование широкой гаммы вариан тов строения венечно-артериальной системы. Однако детально проанализировав вари анты отхождения, соединения и ветвления ВА в более чем 500 сердцах, мы выявили, что в принципе их можно сгруппировать в 11 однотипных групп. Количество сердец в этих группах разнилось: в одних из них оказалось более 100 сердец, в других - 10-20,
85