

Работа электрических ионных каналов
Активация и инактивация электрических ионных каналов зависит от разности потенциалов на мембране, ее деполяризации и реполяризации. Разность потенциалов определяет состояние заслонок, находящихся в канальных порах. Открытие заслонок делает канал проходимым для ионов (активированным), закрытие – не проходимым для них (инактивированным).
Каналы для разных ионов имеют различное количество заслонок. В калиевом канале – одна заслонка, открывающая и закрывающая канал, в натриевом – две: наружная, открывающая канал (активирующая), и внутренняя, закрывающая (инактивирующая). В кальциевом и хлорном каналах также две заслонки.
При рассмотрении механизма возбуждения нас будет интересовать работа главным образом натриевых и калиевых каналов. В покоящейся клетке открыты калиевые каналы и закрыты натриевые. У натриевого канала закрыта наружная (активирующая) заслонка и открыта внутренняя (инактивирующая). Такое положение заслонок зависит от величины электрического тока, текущего через мембрану клетки, находящейся в состоянии покоя. Этот ток называется током покоя. Именно он не дает клетке возбудиться, сохраняет ее стабильное состояние и позволяет осуществлять нормальные метаболические процессы. Для регистрации тока покоя один из микроэлектродов вводят внутрь клетки и как бы касаются им внутренней поверхности мембраны, а другой фиксируют на ее наружной поверхности. Электроизмерительный прибор, включенный между электродами, покажет, что на мембране существует разность потенциалов и через нее течет электрический ток, напряжение которого для большинства клеток равно 70 мВ. При этом внутренняя поверхность мембраны имеет отрицательный заряд, а наружная – положительный.
Не трудно убедиться, что этот ток создается ионами калия. Достаточно заблокировать тетраэтиламмонием (специфический блокатор) калиевые каналы, как ток исчезает. Такой же эффект можно получить, повысив концентрацию калия снаружи клетки и снизив таким путем калиевый концентрационный градиент.
Ионы калия, концентрация которых в клетке значительно выше, чем во внеклеточной жидкости, подчиняясь концентрационному градиенту, стремятся выйти наружу, следующие же за ними анионы органических веществ будут вынуждены «осесть» на внутренней поверхности мембраны. В результате некоторое количество катионов и анионов окажется разобщенным, а мембрана – поляризована. Снаружи она приобретет положительный потенциал, а внутри отрицательный. При наличии открытых калиевых каналов через мембрану потечет электрический ток, который мы ранее обозначили как ток покоя. Ионы натрия и хлора, находящиеся снаружи в высокой концентрации, будут стремиться проникнуть внутрь клетки, однако это удастся только ионам хлора, поскольку натриевые каналы закрыты. \проникая внутрь, ионы хлора будут увеличивать отрицательный потенциал, оставшиеся же снаружи ионы натрия, вместе с вышедшими ионами калия – положительный потенциал.
На образование электрического двойного слоя используется очень мало ионов, поэтому их концентрационные градиенты практически остаются неизменными. По мере того как силы концентрационных градиентов уравновешиваются электростатическими силами, на мембране устанавливается динамическое равновесие. Уровень, на котором установится это равновесие, будет определяться соотношением концентрации ионов по обе стороны мембраны и величиной разности потенциалов на ней.
Трансмембранный потенциал, уравновешивающий диффузию данного иона, называется его равновесным потенциалом и обозначается буквой Е. Например, равновесный потенциал калия в скелетной мышце, у которой калия внутри клеток в 39 раз больше, чем снаружи, будет равен – 95 мВ. Кратко это можно выразить таким образом: К+н : К+в = 1 : 39, соответственно Ек = - 95 мВ.
Все изложенные закономерности могут быть представлены в виде простых физических формул, которые делают их значительно понятнее. Например, мы установили, что ионы К+ испытывают действие двух сил, являющихся следствием концентрационной и электрической потенциальной энергии.
Состояние динамического равновесия перемещения ионов достигается тогда, когда количество концентрационной потенциальной энергии, приходящейся на 1 моль иона, оказывается равным количеству потенциальной электрической энергии, приходящейся на такое же количество ионов.
Потенциальная концентрационная энергия может быть измерена работой, необходимой для переноса через мембрану 1 моля иона при условии неизменности его концентрации. Обозначим эту работу через Wc и в качестве переносимого иона возьмем К+. Расчет показывает, что
Wc
=
RT
ln
[K+]в
- RT
ln
[K+]н
= RT
(ln
[K+]в
- - RT
ln
[K+]н)
= RT
ln
![]() Здесь
R
–
газовая константа, то есть кинетическая
энергия 1 моля вещества при 10
К (8,2
джоуля), а Т
– абсолютная температура. Соответственно
RT
– кинетическая энергия моля ионов при
температуре измерения.
Здесь
R
–
газовая константа, то есть кинетическая
энергия 1 моля вещества при 10
К (8,2
джоуля), а Т
– абсолютная температура. Соответственно
RT
– кинетическая энергия моля ионов при
температуре измерения.
Таким же образом может быть измерена потенциальная электрическая энергия. Она будет соответствовать работе по переносу против электростатических сил моля ионов при условии, что разность потенциалов на мембране остается стабильной. Обозначим эту работу через Wе, она будет равна We = n F Eм, в данной формуле n – валентность иона (для К+ равная 1), F – число Фарадея, то есть заряд моля одновалентных ионов (96500 кулонов), а Eм – электрический потенциал на мембране.
Электрохимический
потенциал может быть представлен как
сумма работ: ∆µ =Wс
+
Wе.
Равновесие достигается, когда ∆µ = 0, то
есть когда Wс
=
- Wе.
Подставляя в это уравнение значение Wс
и
Wе
и заменяя Ем
на Ек
(поскольку рассматриваются условия
равновесия для К+),
находим: RT
ln
[K+]в
= - n
F Eк,
откуда:
EK=-
![]() ln
ln
![]() или EK=
ln
последнее
выражение носит название уравнения
Нернста.
или EK=
ln
последнее
выражение носит название уравнения
Нернста.
Если принять, что Т = 3100 К (370С), и заменить натуральный логарифм на десятичный, то Ек = 60 lg для условий, когда [K+]н : [K+]в = 1 : 39. Ек будет равно – 95 мВ. Для мышечной ткани этот расчетный потенциал почти соответствует истинному – 90 мВ.
Такой же расчет можно сделать для ионов хлора и натрия. В первом случае равновесный потенциал будет близок к калиевому и равен – 90 мВ, во втором – значительно отличаться от него и равняться + 65 мВ. Однако в покое натриевый ток себя не проявляет, так как натриевые каналы инактивированы и мембрана для натрия непроницаема.
Может возникнуть вопрос: почему расчетный ток покоя больше того, который обнаруживается в опыте? Объясняется это некоторой утечкой натрия (проникновением в клетку) через дефекты мембраны и немногие открытые на ней натриевые каналы.
У клеток разных тканей величина тока покоя различна. Это во многом зависит от проникновения в цитоплазму натрия. В нервном волокне ток покоя равен – 70 мВ, а в мышечном – 90 мВ, и всегда ток покоя тщательно охраняется клеткой.
В чем же причина столь бережного отношения клетки к стабильности тока покоя на мембране? Оказывается, от его величины зависит ее функциональное состояние. При деполяризации мембраны возникает возбуждение клетки, при гиперполяризации, наоборот – снижение возбудимости, вплоть до полного торможения.
Чтобы возбудить клетку, надо деполяризовать ее мембрану, то есть уменьшить величину тока покоя. Когда напряжение ока покоя снижается до – 50 мВ, активируются калиевые и натриевые каналы, открываются дополнительные калиевые каналы и множество ранее закрытых натриевых каналов. Скорость их активации различна, калиевые анналы активируются значительно медленнее, чем натриевые. В начале действия раздражителя, когда он еще имеет подпороговую величину, открывается небольшое количество натриевых каналов. Положительно заряженные катионы натрия поступают в клетку, нейтрализуют находящиеся в ней анионы и вызывают снижение разности потенциалов на клеточной мембране. Это снижение носит название местного процесса или местного потенциала. Особенностью местного потенциала является зависимость от силы раздражения. И чем оно сильнее, тем значительнее деполяризация мембраны.
Такая зависимость возникает потому, что чем сильнее раздражение, тем больше открывается натриевых каналов и в клетку входит более количество натрия, нейтрализующего анионы. Местный процесс возникает там, где действует раздражитель и не распространяется по мембране.(стиль)
Как только сила раздражителя достигает пороговой величины, то есть такой, при которой напряжение тока покоя уменьшается до критического значения (обычно до – 50 мВ), активируются быстродействующие натриевые каналы и более медленно действующие калиевые (рис. 3).
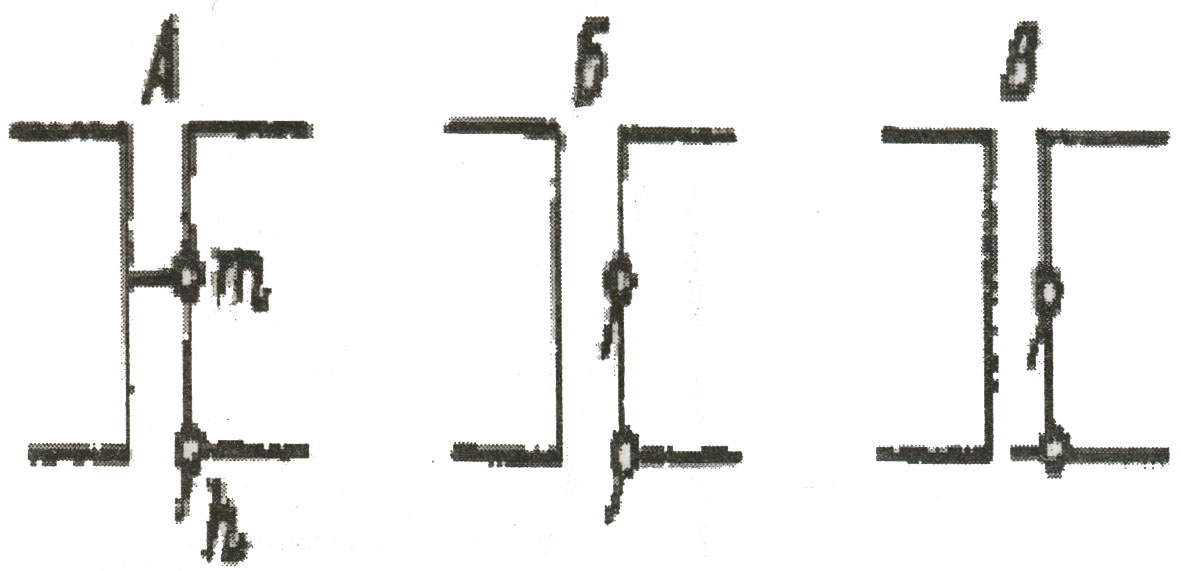
Рис. 3. Схема положения активационных (m) и инактивационных (h) заслонок натриевых каналов: А – в покое, Б – возбуждение, В – инактивированное состояние
Натриевые каналы активируются и инактивируются в течение 1 – 1,5 мс, калиевые несколько отстают от них. За это короткое время через каждый натриевый канал в клетку успевает проникнуть всего лишь 15 – 20 положительных ионов натрия, но, так как каналов очень много (400 на 1 мкм2), происходит деполяризация мембраны до нуля и ее перезарядка. В результате наружная поверхность мембраны становится отрицательной, а внутренняя – положительной.
Этот скачок тока составляет +40 мВ и называется овершут, что в переводе с английского означает перелет. Если сложить отрицательную часть тока на мембране с положительным скачком, получится суммарная амплитуда напряжения тока, равная 110 мВ, это и будет величина деполяризационной фазы тока, который носит название тока действия (биотока) (рис. 4).

Рис. 4. Потенциал действия (биоток, или ток действия). Фаза нарастания, или деполяризации, зависит от вхождения в клетку натрия. Порог, или критический уровень деполяризации мембраны. Фаза реполяризации, зависит от выхода из клетки калия. Фаза гиперполяризации (следовой потенциал)
Ток действия, в сущности, и есть сам процесс возбуждения. Возникнув в нерве, он будет с определенной скоростью распространяться по его длине. В мышечных клетках ток действия запустит сложную цепь биохимических реакций, которые приведут к сокращению. В железах те же изменения вызовут секрецию.
В состоянии покоя в натриевом канале внутренняя заслонка (инактивирующая) открыта, а наружная (активирующая) закрыта (см. рис. 3). В момент, когда мембрана деполяризуется до критического уровня, наружная заслонка открывается, а ионы поступают в клетку. Процесс продолжается до начала фазы реполяризации, которая индуцируется активными калиевыми каналами. Как только разность потенциалов восстанавливается до –20 мВ, внутренняя заслонка закрывается и натриевый канал инактивируется. Спустя некоторое время закрывается и наружная заслонка. Восстановление же нормального тока покоя на мембране является сигналом для открытия внутренней заслонки. Благодаря работе ионных каналов на кривой тока действия отмечается ряд фаз (см. Рис. 4). В момент действия раздражителя возникает местный потенциал, по достижении критической деполяризации развивается деполяризационная фаза, которая сменяется реполяризационной, восстанавливающей нормальный ток покоя. Фаза деполяризации и перезарядки мембраны обязана своим возникновением активации натриевых каналов, фаза реполяризации – активности калиевых. Во время реполяризации клетка испытывает состояние повышенной и пониженной возбудимости.
Повышенная возбудимость объясняется частичной деполяризацией мембраны, благодаря чему раздражителю легче деполяризовать ее до критического уровня. В фазе пониженной возбудимости, которую называют также положительным шлейфом биотока, или андершутом, что в переводе с английского означает недолет, мембрана имеет разность потенциалов на 5 – 10 мВ больше значения тока покоя, то есть гиперполяризована. Соответственно достижение критического уровня в этом случае делается затруднительным.
Природа этих изменений не очень ясна. Полагают, что фаза повышенной возбудимости, являющаяся по существу замедлением реполяризации, связана с остаточной активностью натриевых каналов. Фазу пониженной возбудимости связывают с работой Na, К – насоса, активированного натрием, накопившимся в клетке за время возбуждения.
При возбуждении происходит изменение возбудимости клетки. В фазу деполяризации, когда активированы натриевые каналы, она абсолютна невозбудима. Этот период называют абсолютной рефрактерной фазой. Когда же происходит реполяризация и натриевые каналы инактивируются, а калиевые восстанавливают нормальный ток покоя, возникает относительная рефрактерная фаза, во время которой возбудимость постепенно возвращается к норме. В конце реполяризации возбудимость делается выше нормы (фаза экзальтации) и далее, при гиперполяризации, вновь снижается.
В нервном волокне абсолютная рефрактерность длится 1 – 1,5 мс, как раз столько времени, сколько активны натриевые каналы. Относительная рефрактерность занимает примерно столько же времени, однако в конце из-за экзальтации этот период несколько затягивается и иногда может длиться до 10 мс.
Фаза гиперполяризации в нервном волокне обычно короткая (5 – 10мс), в нейроне же может достигать 50 и более мс.
Ток действия является тем механизмом, с помощью которого кодируется и передается вся информация в рефлекторной дуге. В нормальных условиях эти сигналы генерируют рецепторы, превращая энергию раздражителя в каскад (поток) биотоков, равных по амплитуде. Информация в этом потоке кодируется путем изменения частоты импульсов и интервала между отдельными их «пучками». Такое кодирование называется частотно-интервальной модуляцией.
Рецепторы, образно говоря, являются «переводчиками», осуществляющими перевод характеристик раздражителя на «язык», понятный центральной нервной системе. Ее нейроны, подвергнув полученную информацию обработке, в виде частотного кода направляют регулирующие сигналы к эффекторам – мышцам и железам.
Чтобы закончить наше знакомство с возбуждение, попытаемся объяснить, каким образом при изменении степени поляризованности мембраны закрываются и открываются заслонки ионных каналов. В настоящее время это можно сделать только гипотетически.
Наиболее вероятным кажется объяснение, что заслонки представляют собой белковые субъединицы, снабженные электрически чувствительными сенсорами, в состав которых входят положительно ионизированные группы. При деполяризации мембраны снижается напряжение ее электрического поля, что приводит к конформации сенсора и связанной с ним субъединицы белка заслонки. Не исключено также, что сам сенсор выполняет одновременно функцию заслонки.
Согласно другой гипотезе ионизированные группы канального белка создают в его просвете электрическое поле определенной напряженности и знака, которое регулирует избирательность и способность канала пропускать ионы. При деполяризации мембраны напряженность поля снижается, и канал активируется, при гиперполяризации напряженность возрастает и канал инактивируется.