

Механизм тонуса скелетных мышц
Как уже упоминалось, в условиях организма мышцы работают в фазическом и тоническом режимах. Фазический режим служит для выполнения целенаправленных движений (сгибание, разгибание конечностей, трудовые процессы и т.п.), тонический – для ориентации тела в пространстве, фиксации суставов при фазическом движении и т.д.
Кроме того, по характеру изменения длины мышцы ее сокращение можно разделить на изотоническое и изометрическое. При изотоническом сокращении изменяется дляна мышцы, при изометрическом – длина остается постоянной, а изменяется напряжение.
В условиях организма редко встречается чисто изометрическое или изотоническое сокращение. Обычно, при работе мышцы изменяется ее длина и напряжение. Поэтому вводят еще один термин, обозначающий естественный режим работы мышц: ауксотонический, или анизотонический, тип сокращения, при котором меняется и длина, и напряжение мышцы.
Изотонический и анизотонический режимы определяют динамическую работу локомоторного аппарата. При динамической работе выделяют еще два типа сокращения мышцы. Первый из них называется концентрический, второй эксцентрический.
В первом случае нагрузка на мышцу такова, что при сокращении она все же может ее преодолеть, укорачиваясь при этом. Во втором случае нагрузка столь велика, что напряжение мышцы не в состоянии ее преодолеть, тогда она удлиняется, совершая отрицательную (уступающую) работу.
Тонический режим имеет большое значение для организма. Нельзя осуществить даже самое простое целенаправленное движение, не изменив при этом тоническое состояние мышц тела. И дело не только в том, что с помощью тонуса осуществляется ориентация тела в пространстве и поддержание равновесия при движении конечностей и головы. Тонус мышц превращает наши суставы из системы в механизм.
Под системой в механике понимают такое устройство, которое позволяет двигать соединенные друг с другом детали во всех плоскостях, т.е. имеет неограниченное число степеней свободы. Механизмом же называют устройство, имеющее только одну степень свободы, позволяющее двигаться только в одной плоскости.
Многие суставы нашего тела обеспечивают движение во многих плоскостях и в то же время, при необходимости, осуществляют целенаправленное фазическое движение в одной плоскости. Таким образом, они могут, по мере надобности, быть и системой и механизмом.
Превращение сустава, осуществляющего движение во многих плоскостях, в такой, которой позволяет двигаться только в одной плоскости, происходит благодаря увеличению тонуса расположенных вокруг него мышц. Это увеличение строго координировано и управляется сложными тоническими рефлексами, о которых речь пойдет далее. Для понимания же механизма тонуса нам, прежде всего, надо будет познакомиться с устройством, которое называется нейромоторная единица.
Нейромоторные (двигательные) единицы
Любая мышцы представляет собой сложный орган, имеющий свой нервный, рецепторный и сосудистый аппарат. Отдельные мышечные волокна соединены друг с другом эндо- и перимизиумом, снаружи мышцы покрыта фасцией и снабжена сухожилиями.
В состав мышцы входят три вида волокон (рис. 19): 1 – медленные, неутомляемые (по старой номенклатуре – красные, тонические волокна), 2 – быстрые, легко утомляемые (по старой номенклатуре – белые волокна) и 3 (переходный тип) – быстрые, устойчивые к утомлению.
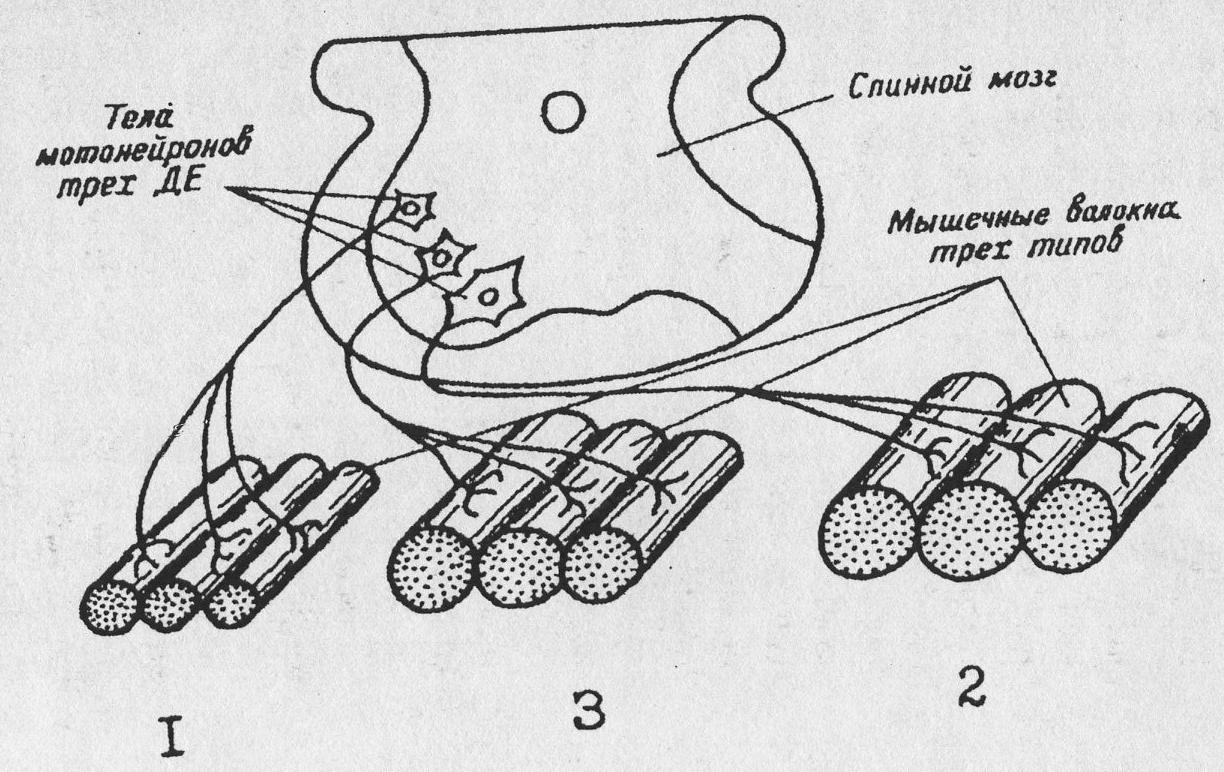
Рис. 19.Схематическое изображение нейромоторных единиц
(двигательных единиц – ДЕ. по Ткаченко): 1. - медленные, неутомляемые, тонические ДЕ, 2 – быстрые, легко утомляемые ДЕ, – быстрые, устойчивые к утомлению ДЕ. Описание в тексте
Отдельные мышечные волокна объединяются в нейромоторные единицы (НЕ), которые иннервируются одним нейроном и функционируют как единое целое.
Нейромоторные единицы 1-й группы иннервируются малыми альфа-мотонейронами, лежащими там же, где и большие альфа-мотонейроны – в передних рогах спинного мозга. Они обладают тонкими аксонами (до 10 мкм), проводящими возбуждение со скоростью 30 м/с. На периферии аксоны делятся на большое число веточек. Каждое мышечное волокно в ДЕ 1-й группы имеет много, иногда десятки нервно-мышечных контактов (синапсов или нервно-мышечных пластинок), разбросанных по всей его длине (на рис. 19 не показано).
Дело в том, что медленные мышечные волокна имеют мембрану, не способную проводить возбуждение. Нервный импульс вызывает в волокнах ДЕ 1-й группы лишь местное возбуждение и сокращение, захватывающее область мионеврального синапса, благодаря же иннервации во многих точках тоническое волокно может сокращаться как единое целое.
Быстрые, фазные ДЕ 2-й группы, приспособленные для выполнения быстрых движений, иннервируются большими альфа-мотонейронами передних рогов спинного мозга, посредством толстых аксонов, имеющих диаметр 10 – 20 мкм и скорость проведения до 120 м/с.
Мышечные волокна фазных ДЕ образуют с нервным волокном 1 – 2 нервно-мышечных синапса. Их возбуждение сопровождается возникновением распространяющегося потенциала действия и сокращением всего волокна, которое протекает быстро, в течение 36 мс.
В фазных ДЕ все приспособлено для быстрого сокращения. Мышечные волокна обладают хорошо развитым миофибриллярным аппаратом, содержащими кальций цистернами, расположенными вблизи миофибрилл, и высокой активностью Са-АТФазного насоса, который возвращает кальций в цистерны (см. первую часть руководства). В то же время они быстро утомляются, так как содержат малое количество миоглобина и хуже кровоснабжаются по сравнению с медленными, тоническими волокнами.
Быстрые, устойчивые к утомлению ДЕ (рис. 19, 3) занимают промежуточное положение. Входящие в их состав мышечные волокна иннервируются так же, как и быстрые, утомляемые. Различие заключается в длительности потенциала действия и скорости сокращения. У быстрых неутомляемых волокон потенциал в два раза длиннее и скорость сокращения в 5 раз меньше, чем у быстрых утомляемых волокон. Кроме того, быстрые неутомляемые волокна содержат больше миоглобина в сравнении с быстрыми утомляемыми.
Для поддержания тонического напряжения наиболее подходящими являются медленные неутомляемые ДЕ. Входящие в их состав волокна не реагируют на одиночный нервный импульс. Для сокращения требуется ряд импульсов (обычно не менее 10), которые поступают с таким интервалом, чтобы обеспечить суммацию локальных потенциалов.
Сокращение тонических волокон развивается очень медленно и потому не требует высокой частоты стимуляции. Малые альфа-мотонейроны, по этой причине, управляют неутомляемыми ДЕ с помощью редких импульсов возбуждения с частотой 4 – 8 имп/с, в то время как большие альфа-мотонейроны регулируют сокращение быстрых ДЕ импульсами с частотой 30 – 5- имп/с.
Степень тонического напряжения мышцы зависит от числа сокращающихся тонических нейромоторных единиц и, следовательно, от числа возбужденных малых альфа-мотонейронов. Варьируя число таких клеток, можно решать сразу две задачи: во-первых, изменять степень тонического напряжения, а во-вторых, делать ДЕ неутомляемыми. Последнее достигается с помощью механизма, меняющего работавшие НЕ на находившиеся в покое.
Механизм чередования нейромоторных единиц основан на наличии в дуге тонических рефлексов тормозных вставочных нейронов (клеток Реншоу) и коллатералей аксонов малых альфа-нейронов, которые, возвращаясь обратно в спинной мозг, контактируют с клетками Реншоу (рис. 11, Б).
Процесс осуществляется следующим образом. Тоническое сокращение возникает в результате стимуляции малых альфа-нейронов проприоцептивными импульсами от рецепторов той же мышцы, т.е. благодаря включению миотатического рефлекса. Раздражителем, удлиняющим мышцу, является ее собственный вес.
Возбужденные мотонейроны тонических нейромоторных единиц направляют импульсы на периферию к соответствующим мышечным воокнами и по коллатералям, в порядке обратной связи, к тормозным клеткам Реншоу. Последние, контактируя с нейронами функционирующих нейромоторных единиц, их тормозят давая «отдохнуть» работавшими мышечным волокнами.
Одновременно под влиянием непрекращающегося потока проприоцептивных импульсов повышается возбудимость нейронов не работавших нейромоторных единиц, которые начинают функционировать, сменяя предыдущие.
Таким образом, благодаря функционированию механизма, «сменяющего» нейромоторные единицы, тоническое напряжение мышцы делается неутомляемым.
В центральной нервной системе тонический режим работы мышц могут создавать все отделы стволовой части, начиная со спинного мозга. Однако последний вызывает очень слабое тоническое напряжение, что объясняется низкой возбудимостью его малых альфа-мотонейронов. Надо иметь ввиду, что эти клетки вообще обладают низкой возбудимостью, а после удаления головного мозга их возбудимость снижается еще более.
Тем не менее, на спинальном препарате лягушки легко убедиться, что спинномозговой тонус все же есть. У такого препарата задние лапки слегка согнуты в коленных суставах, и если на одной стороне перерезать седалищный нерв, лапка этой стороны повиснет «плетью». Следовательно, ее мышцы находились в слабом тоническом напряжении.
Тонус, создаваемый спинным мозгом, не позволяет животному даже стоять на конечностях, однако он является основой механизма, дальнейшее усовершенствование которого дает возможность осуществлять все разнообразие тонических рефлексов. Для усиления тонических рефлексов спинного мозга требуется «вмешательство» стволовых центров, к описанию которых мы далее и переходим.
Двигательная функция ствола головного мозга
В стволовой части головного мозга, которая включает в себя продолговатый мозг, мост и средний мозг, сосредоточены основные ядра, обеспечивающие выполнение статических и статокинетических тонических рефлексов (рис. 20).

Рис. 20.Локализация двигательных центров в стволе мозга (по Шмидту).
Описание в тексте
На границе между продолговатым мозгом и мостом расположены 4 вестибулярных ядра (на рис. 20 показано одно ядро), среди которых главную роль играет ядро Дейтерса. Эти ядра наряду с другими ядрами ствола обеспечивают выполнение статических рефлексов позы, или положения.
Отростки нейронов вестибулярного ядра опускаются в спинной мозг, образуя медиальный вестибулоспинальный тракт (рис. 21). Через вставочные нейроны волокна тракта усиливают тонус разгибателей и тормозят тонус сгибателей. Торможение, пост- и пресинаптическое, осуществляется интернейронами с помощью гамма-аминомаслянной кислоты и глицина. Эти медиаторы активируют хлорные каналы. При постсинаптическом торможении они гиперполяризуют мембрану альфа- и гамма-мотонейронов, а при пресинаптическом торможении деполяризуют пресинаптические нервные окончания.

Рис. 21. Поперечный срез спинного мозга с обозначение проводящих путей стволовых ядер (по Шмидту). Описание в тексте
В регуляции тонуса кроме вестибулярных ядер принимает участие ретикулярная формация ствола. В области варолиева моста локализовано ядро (рис. 20), нейроны которого своими отростками, входящими в состав ретикулоспинального медиального тракта (рис. 21), подобно нейронам вестибулярных ядер, тормозят сгибатели и стимулируют разгибатели. Механизм действия аналогичен вышеописанному.
Противоположным действием обладает ретикулярная формация продолговатого мозга (рис. 20). Ее нейроны образуют в спинном мозге латеральный ретикулоспинальный тракт (рис. 21), который через вставочные нейроны, используя механизмы, рассмотренные выше, тормозит разгибатели и стимулирует сгибатели.
Децеребрационная ригидность
Если отделить средний мозг от моста и продолговатого мозга, т.е. произвести перерезку ствола мозга по линии 1 (рис. 20), возникает децеребрационная ригидность (рис. 22), резко увеличивается тонус во всех мышцах, но поскольку разгибатели сильнее сгибателей, животное принимает разгибательное положение. При этом все четыре конечности будут максимально разогнуты, спина выгнута, а голова и хвост запрокинуты к спине.

Рис.22. Кошка в состоянии децеребрационной ригидности. Описание в тексте
Децеребрационная ригидность может быть обусловлена двумя механизмами и соответственно называется α- и γ-ригидность. В основе гамма-ригидности лежит рефлекторный процесс, который обусловлен резким возрастание возбудимости гамма-петель и альфа-мотонейронов, особенно разгибателей. В результате проприоцептивные импульсы вовлекают в реакцию почти все нейромоторные единицы.
В рефлекторной природе этого вида ригидности легко убедиться, если попытаться согнуть разогнутые лапы, они оказывают при этом активное сопротивление в связи с растяжением разгибателей и увеличением проприоцептивной импульсации.
Еще более убедительно о рефлекторной природе гамма-ригидности свидетельствует опыт с перерезкой задних корешков на одной из конечностей. При этом ригидность на ней немедленно исчезает. Деафферентация конечности прерывает поток импульсов от проприорецепторов мышц и исключает проявление децеребрационной ригидности.
Альфа-ригидность проявляется при ишемической децеребрации, возникающей в результате перевязки сонных и основной артерий. Такая операция выключает большую часть мозжечка и моста, что приводит к повышению возбудимости альфа-мотонейронов спинного мозга и увеличению их реакции на импульсы из ядра Дейтерса. Поток последних увеличивается, поскольку ослабевает тормозное влияние мозжечка.
Гамма-децеребрационную ригидность следует отнести к позным тоническим рефлексам, которые обеспечивают животному основную стойку, возможность стоять на четырех конечностях и противодействовать силе тяжести.
Чтобы перестраивать тонус мышц при изменении положения тела, т.е. осуществлять тонические выпрямительные рефлексы, необходимо сохранение среднего мозга. В среднем мозге локализованы нейроны красного ядра (рис. 20), которые получают информацию от всех отделов центральной нервной системы, участвующих в регуляции локомоции.
С помощью руброспинального тракта (рис. 21), через вставочные нейроны, красное ядро устраняет децеребрационную ригидность, ослабляя тонус разгибателей и усиливая тонус сгибателей, в результате животное получает возможность перестраивать тонус при изменении положения тела.
Мезэнцефальное животное в отношении локомоции мало чем отличается от нормального. В этих условиях источником информации для осуществления тонических рефлексов являются вестибулярный аппарат, проприорецепторы шеи и экстерорецепторы кожи.
Для анализа роли отдельных видов рецепторов в выпрямительных рефлексах производят их последовательное выключение. Чтобы выяснить значение вестибулярного аппарата, посредством деафферентации, наложения гипсового ошейника или новокаиновой блокады, выключают проприорецепторы шеи.
При вращении такого животного вокруг оси, проходящей через оба виска, наблюдается увеличение тонуса разгибателей всех конечностей, когда оно находится в положении – затылком к земле, и сгибание их при нормальном положении – спиной вверх. В данном случае относительное положение всех частей тела оставалось неизменным, менялось только направление силы тяжести по отношению к телу.
Соответственно, тонический рефлекс мог проявиться только при раздражении тех рецепторов, которые чувствительны к изменению направления земного притяжения, т.е. вестибулярных аппаратов.
У животных с разрушенными лабиринтами можно получить выпрямительный рефлекс, если изменить положение головы по отношению к туловищу. При этом происходит раздражение проприоцепторов шейных мышц и изменение тонуса в различных мышечных группах. Следует заметить, что для такой реакции необязательно сохранение среднего мозга. Выпрямительная реакция сохраняется и у децеребрированных животных, проявляясь даже лучше, чем у интактных, у которых импульсы вышележащих центров осложняют картину.
Запрокидывание головы вверх вызывает повышение тонуса разгибателей передних конечностей и снижение тонуса разгибателей задних. В результате животное, приседая на задние конечности, сохраняет равновесное положение тела (рис. 23, А). Наклон головы вниз (рис. 23, Б) вызывает противоположную реакцию: снижение тонуса разгибателей передних конечностей и увеличение тонуса разгибателей задних.

Рис. 23. Изменение тонуса мышц конечностей у децеребрированной кошки с разрушенными вестибулярными аппаратами: А – при запрокидывании головы вверх, Б – при наклоне головы вниз
Поворот головы вправо и влево также приводят к изменению тонуса мышц конечностей. Поворот головы вправо увеличивает тонус разгибателей правой передней конечности и сгибание левой. Поворот головы влево вызывает противоположную реакцию.
Для проявления тонических выпрямительных рефлексов в физиологических условиях нужно участие вестибулярных аппаратов, проприорецепторов шеи и всех мышц тела, а также рецепторов кожи.
После операции перерезки мозга выше четверохолмия (см. рис. 20, 2) животное, спустя некоторое время, поднимает голову, затем устанавливает все туловище в правильное положение, становится на ноги и принимает естественную позу.
В положении лежа на боку раздражаются рецепторы кожи одной стороны туловища и вестибулярный аппарат. Это вызывает подъем и правильную установку головы. Изменение положения головы неравномерно растягивает мышцы шеи, раздражая локализованные в них проприорецепторы. Чтобы устранить деформацию мышц шеи, необходимо повернуть туловище, в результате все тело животного примет нормальное положение.
Для включения выпрямительного тонического рефлекса достаточно раздражения кожных рецепторов. Если положить на бок животное с разрушенным вестибулярным аппаратом, оно немедленно поднимает голову и устанавливает е в правильное положение – затылком вверх. Реакция отсутствует, если сверху поместить легкую дощечку и таким образом с двух сторон уравновесить раздражение кожных рецепторов.
Этот сложный тонический рефлекс развивается в строгой последовательности. В физиологических условиях, в первую фазу реакции, нормальное положение занимает только голова. Далее, во вторую фазу, раздражаются проприорецепторы шеи и рецепторы кожи. Чтобы устранить это раздражение, животное вынуждено посредством изменения тонуса мышц тела установить его в правильное положение.
У мезенцефальных животных наблюдается еще один вид тонических рефлексов, которые называются статокинетические. Эти рефлексы возникают при угловом и линейном ускорении. Информацию для их проявления поставляют зрительные и вестибулярные рецепторы.
При вращении тела голова сначала медленно отклоняется в сторону, противоположную вращению, затем, быстрым движение, возвращается в нормальное положении.
Такое же наблюдается при движении глазных яблок и носит название нистагм. Различают медленный и быстрый компоненты нистагма. Вначале глазные яблоки медленно отклоняются в направлении, противоположном вращению (медленная компонента нистагма), а затем быстро возвращаются в исходное положение (быстрая компонента). Реакция зависит от вестибулярного аппарата и может служить тестом для определения чувствительности его полукружных каналов.
К статокинетическим рефлексам относится также «лифтная» реакция, которая проявляется при линейном ускорении, например, при подъеме и опускании площадки, на которой сидит животное. Рефлекс выражается в увеличении тонуса разгибателей конечностей при движении площадки вверх и увеличении тонуса сгибателей при движении вниз.