

Двигательная функция коры головного мозга
В этом разделе мы познакомимся с устройством и функцией двигательной области коры головного мозга, более подробное описание корковой функции будет дано в третьей части пособия, посвященной высшей нервной деятельности.
Двигательная кора (первичная моторная область) находится в прецентральной извилине цитоархитектонического поля 4 по Бродману (рис. 29).
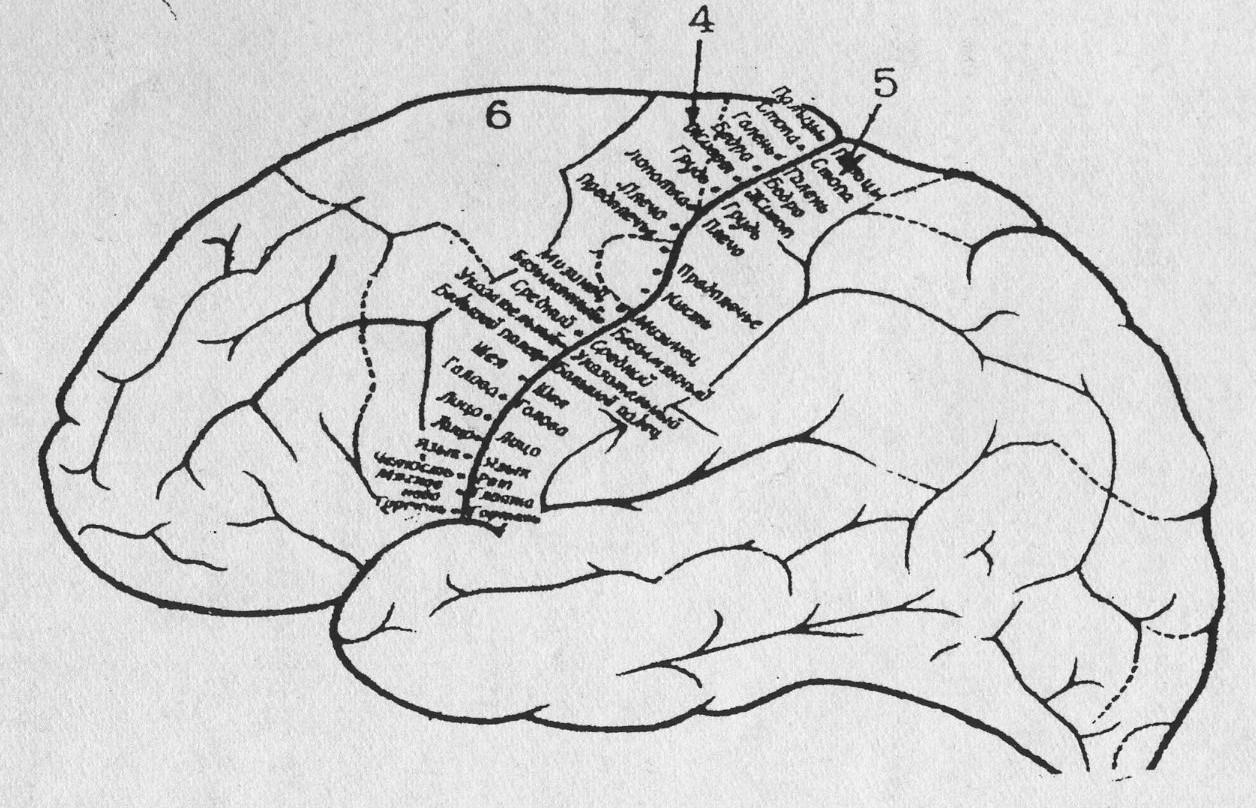
Рис. 29. Схема моторной коры с обозначением полей по Бродману:
4– передняя центральная извилина - двигательная, 5 – задняя центральная извилина – сенсорная, 6 – вторичная моторная область. Описание в тексте
В этом районе кора имеет большую толщину и в ней отсутствует внутренний зернистый слой (рис. 30, слой IV). В пятом слое (рис. 30) локализованы гигантские пирамидные клетки Беца, поэтому эта область называется агранулярная гигантопирамидная область.

Рис. 30. Схематическое изображение шестислойной коры:
I – молекулярный слой, образован, в основном. Сплетением нервных волокон. II – наружный зернистый слой. В нем расположены вставочные нейроны, имеющие форму круглых, треугольных и многоугольных зерен. III - слой малых пирамид. IV – внутренний зернистый слой. Подобно второму
слою, состоит из мелких вставочных нейронов (отсутствуют в моторной области коры). V – слой больших пирамид Беца. От верхней части этих клеток отходит толстый дендрит, вертикально поднимающийся вверх и многократно ветвящийся в поверхностном слое коры. VI – мультиформный слой, состоит из нейронов треугольной и веретенообразной формы
Наряду с полем 4, двигательные центры локализованы во вторичной моторной области (рис.29, поле 6). Которая переходит на медиальную поверхность коры (на рис. 29 показана частично), включая в себя дополнительную моторную область и латеральную премоторную кору. При достаточно длительном раздражении поля 6 возникает вращение туловища и глаз с подниманием контралатеральной руки. Столь сложная реакция объясняется процессами суммации в многочисленных нейронных цепях.
Установлено, что в передней центральной извилине, имеются неравномерно распределенные зоны, управляющие различными мышцами тела. Наибольшее место занимает представительство мышц лица, языка, кистей рук, т.е. тех частей тела, которые несут наибольшую функциональную нагрузку. В то же время сравнительно мало представлены мышцы туловища и ног (рис. 31).

Рис. 31.Соматотопическая карта, показывающая влияние раздражения
отдельных точек прецентральной извилина на сокращение мышц (двигательный гомункулус). Описание в тексте
Несмотря на сложное цитологическое строение моторной коры, все нейроны, входящие в ее состав, можно разделить на две группы: пирамидные и звездчатые клетки (рис. 30, IV слой). Большие и малые пирамиды являются основными, командными нейронами коры. Их аксоны образуют пирамидный тракт – единственный эфферентный канал, по которому импульсы проводятся ко всем нижележащим центрам. Звездчатые клетки выполняют функцию вставочных нейронов, обрабатывающих афферентную информацию и передающих сигналы пирамидным клеткам и в другие центры коры.
Благодаря особой форме пирамидных клеток, имеющих треугольное тело, вертикально поднимающийся вверх дендрит, который проходит через все слои коры и систему горизонтальных дендритов, создаются условия для возникновения оригинальной корковой нейронной архитектуры.
Окружающие пирамидную клетку звездчатые вставочные нейроны образуют с ее вертикальным дендритом до 4000 синапсов, для чего на дендрите имеются многочисленные шипики. В результате создается колонка-модуль, весьма удобное устройство для обработки информации.
Вокруг каждой пирамидной клетки выстраивается микроколонка, имеющая диаметр около 100 мкм. Микроколонки (микромодули) объединяются друг с другом, образуя макромодули, и так далее. На периферии один микромодуль управляет группой нейромоторных единиц мышцы, макромодуль может управлять уже всей мышцей, а объединение макромодулей – несколькими мышцами и, наконец, всей конечностью.
Следует также учитывать, что влияние вставочных нейронов на вертикальный и горизонтальный дендриты пирамидной клетки различно. При воздействии на вертикальный дендрит возникает деполяризация мембраны и возбуждение пирамидной клетки, воздействие же на горизонтальные дендриты вызывает гиперполяризацию мембраны и торможение нейрона. Различие в действии вставочных нейронов расширяет возможности модуля по обработке информации.
Пирамидный тракт, состоящий из аксонов больших и малых пирамидных клеток, включает в себя миелинизированные и немиелинизированные волокна. Примерно 70 % тракта приходится на аксоны малых пирамид и 30 % составляют аксоны больших пирамид Беца. Среди 70 % как раз и встречаются не покрытые миелином, безмякотные волокна, пирамиды же Беца имеют аксоны с диаметром 12–22 мкм, покрытые миелином и проводящие возбуждение со скоростью до 120 м/с.
Входящие в состав пирамидного тракта аксоны различаются и в функциональном отношении. Принадлежащие пирамидам Беца нервные проводники проходят в спинной мозг, где оканчиваются на вставочном нейроне или прямо на альфа-мотонейроне. По этой причине раздражение больших пирамид вызывает, через короткий латентный период, локальную двигательную реакцию.
Малые пирамиды локализованы не только в поле 4, но и в поле 6 по Бродману (рис. 29). Их аксоны, составляющие большую часть пирамидного тракта, по дороге (пути) к спинному мозгу отдают многочисленные коллатерали нижерасположенным центрам, участвующим в регуляции движения.
Коллатерали направляются к полосатому телу и через него – к таламусу, мозжечку, красному ядру, двигательным и тоническим центрам моста и продолговатого мозга. После частичного перекреста, вместе с толстыми аксонами пирамид Беца, аксоны малых пирамид замыкаются на вставочных нейронах той же и противоположной сторон спинного мозга.
Столь сложный путь, с вовлечением в реакцию многочисленных центров, совокупность которых называется экстрапирамидной системой, не только удлиняет латентный период, но и вызывает, вместо локальных движений, сложные, сочетанные двигательные реакции, характерные для раздражения поля 6.
Вначале, когда впервые были открыты двигательные зоны коры, полагали, что здесь и создаются сложные двигательные, волевые программы. Позднее пришли к выводу, что такие программы создаются хотя и с участием этих зон, но главным образом в ассоциативных полях коры лобной, височной, теменной областей. Соответственно, двигательной области коры отвели более скромную роль – исполнительного звена локомоторной системы.
Кроме эфферентных, моторная кора имеет обширные афферентные связи. Источниками информации являются задняя центральная извилина и ядра таламуса, в которых проецируются все виды мышечно-суставной и кожной чувствительности.
Информацию передают вставочные нейроны, звездчатые клетки, клетки-зерна II слоя и веретенообразные клетки VI слоя коры. Нейроны VI слоя своими отростками создают тесные связи с таламусом, образуя центрофугальные и центропитальные таламокортикальные тракты.
Через таламус в кору больших полушарий, наряду с обширным потоком информации от двигательных центров, поступают модулирующие импульсы ретикулярной формации, без активирующего влияния которых кора не может функционировать.
Таким образом, таламокортикальные связи являются важнейшими входными (афферентными) «воротами» коры. Говоря образно, таламус представляет собой своего рода «референтный центр» по предварительной обработке и отбору жизненно важной для организма информации. Перед тем как она направляется в кору головного мозга.