

Я.Д. Финкинштейн общая физиология центральной нервной системы новосибирск
2001
Введение
Наверное, нет необходимости говорить о важности знаний по физиологии центральной нервной системы при изучении психологии. Однако, учитывая трудность стоящей перед студентами задачи, следует хотя бы кратко сказать о том, какую роль играют физиологические закономерности в психике человека.
Уже одно перечисление тех проблем, которыми нам предстоит заниматься, убедительно показывает насколько важно заложить в основу свих психологических знаний представление о физиологических процессах.
Наш курс начинается с изучения рефлекторной реакции, того основного механизма, который центральная нервная система использует в своей деятельности. Эта реакция протекает на базе анатомического субстрата, именуемого рефлекторной дугой, или кольцом (если учитывается обратная связь). Центральная нервная система является коллектором огромного количества рефлекторных дуг.
Во многих случаях к рефлекторному механизму подключается одна из эндокринных желез, вырабатывающих гормоны. И тогда регуляторные возможности рефлекса значительно расширяются.
В первой книге методического пособия мы совершим «путешествие» по рефлекторной дуге и прежде всего, изучим механизм возбуждения, который, без преувеличения, может считаться основным физиологическим процессом.
Все звенья рефлекторной дуги функционируют благодаря способности развивать возбуждение. Любой раздражитель, действующий на организм, посредством рецепторов рефлекторной дуги превращается в импульсы возбуждения, которые подвергаются обработке в центральной нервной системе. Образно говоря, импульсы – это единственный «язык», который она понимает.
Благодаря возбуждению функционируют нейроны нервных центров, производя обработку импульсного кода. Возбуждение обеспечивает передачу командных сигналов эффекторам – мышцам и железам, регулируя их деятельность.
Мы познакомимся с законами проведения возбуждения в нервных проводниках и синапсах – местах контактов между отдельными элементами рефлекторной дуги и, наконец, рассмотрим, как функционируют эффекторы, взяв в качестве примера скелетные мышцы.
Во второй книге пособия мы перейдем к изучению механизмов взаимодействия между рефлекторными дугами, чтобы разобраться в двигательных реакциях организма. Здесь уместно будет вспомнить высказывание И.М. Сеченова о том, что вся психология человека выражается в его действиях, те есть в двигательных реакциях. В этих реакциях невозможно разобраться без изучения интегративной функции центральной нервной системы, взаимодействия между ее центрами.
Надеюсь, я убедил Вас в полезности тех знаний, которые Вы приобретете, ознакомившись с данным методическим руководством. В дальнейшем они Вам, несомненно, помогут при изучении психологии.
Рефлекторная регуляция и механизм возбуждения
В этом разделе мы познакомимся с механизмами регуляции, которые свойственны всем системам организма, а затем займемся изучением процесса возбуждения, являющегося основной физиологической реакцией, оживляющей работу его органов.
Регулирующие механизмы всех систем организма построены по типу рефлекторной реакции. Впервые понятие рефлекса было введено в физиологию Декартом в XVII веке. Под рефлексом понималась отраженная реакция, протекающая при обязательном участии центральной нервной системы. Последнее надо хорошо запомнить. Потому что в организме есть немало реакций, сходных с рефлекторной, которые протекают на периферии, без непосредственного участия центральной нервной системы.
Как правило, любая схема (система регуляции) регуляторного механизма состоит из четырех звеньев: информационного, центрального, исполнительного и обратной связи. Информационное звено поставляет в систему данные о регулируемой реакции, центральное – обрабатывает эти данные, исполнительное – осуществляет саму регуляторную операцию, а обратная связь, которая бывает отрицательной и положительной, т. е. тормозящей и стимулирующей, действует на центр, корригируя процесс регулирования.
Все эти звенья мы обнаруживаем и в физиологических регуляторных системах, где они входят в состав рефлекторной дуги, или кольца (если учитывать обратную связь). Информационное звено представлено в рефлекторной дуге рецепторами, центральное – центрами центральной нервной системы, исполнительное – эффекторами, различными видами мышечной и железистой ткани, выполняющими механическую и химическую функцию (рис. 1).

Рис.1. Рефлекторная дуга: 1 – рецептор, 2 – чувствительный нейрон (афферентный), 3, 4 – вставочные нейроны, 5 – двигательный нейрон (эфферентный), 6 – скелетная мышца (эффектор).
Кроме рефлекторных дуг, состоящих только из нервных элементов, существуют и такие, которые включают в свой состав эндокринные железы. Эти механизмы называются рефлексами с гуморальным или гормональным звеном, и на их основе строится нейрогормональная регуляция.
Отдельные звенья рефлекторной дуги соединяются нервными проводниками периферической нервной системы. Те из них, которые проводят импульсы возбуждения от рецепторов в центральную нервную систему, называются афферентными, те же, которые проводят импульсные сигналы к эффекторам – эфферентными. В рефлекторных дугах с гормональным звеном в эфферентную часть включается эндокринная железа, которую можно назвать эффектором первого порядка. Соответственно эффектором второго порядка будет орган-мишень.
При включении рефлекторного механизма происходит не просто запуск эффектора, а приспособление его работы к запросам организма. Это значит, что рефлекторная реакция целесообразна и соответствует характеристикам вызвавшего ее раздражителя.
В свое время И. П. Павловым были сформулированы три основных принципа рефлекторной теории, позволяющих оценить биологическое значение рефлекторной регуляции. Первым был назван принцип детерминированности, то есть причинной обусловленности реакции. Согласно этому принципу, любая рефлекторная реакция причинно обусловлена и возникает только под влиянием раздражителя. В соответствии со вторым принципом при осуществлении рефлекторной реакции происходит анализ и синтез раздражителя, позволяющий сделать реакцию целесообразной для организма.
Третий принцип, структурности, постулирует невозможность проявления рефлекторной реакции при нарушении функциональной целости рефлекторной дуги, то есть физиологической, а не анатомической целости, последняя сохраняется и у трупа.
Мы познакомились с механизмами, осуществляющими регуляцию в любой физиологической системе. (К более подробному рассмотрению, изучению мы в главе…) Более подробное их рассмотрение будет сделано в разделе о центральной нервной системе, здесь же уместно перейти к описанию механизма возбуждения.
Возбуждение, без преувеличения, можно назвать основным физиологическим процессом. В сущности, и сама физиология может быть определена как наука о возникновении и превращении возбуждения в органах и системах организма. В самом общем виде возбуждение можно определить как реакцию на раздражение физиологической системы, изменяющее ее функцию. В физиологии возбудимыми принять считать органы и ткани, отвечающие на раздражение специфической реакцией. К ним относятся мышечная, нервная и железистая ткани.
Возбуждение можно изучать на любом возбудимом объекте, однако удобнее всего это делать на нервно-мышечном препарате, изготовленном из задних лапок лягушки. Препарат этот впервые был предложен в XVII веке Сваммердамом и вот уже в течение трех веков добросовестно служит как для физиологических исследований, так и для обучения студентов.
Нервно-мышечный препарат состоит из двух классических возбудимых тканей: нервной и мышечной. По сокращению мышцы можно наблюдать за появлением возбудительного процесса в нерве. Кроме того, препарат весьма неприхотлив. Для поддержания нормального функционального состояния требуется лишь периодическое (орошение) смачивание его физиологическим раствором (раствором Рингера), имитирующим солевой состав плазмы крови лягушки.
Возбуждение нервно-мышечного препарата может быть вызвано любым раздражителем. Например, механическим, термическим, химическим, электрическим. Однако из этого набора один вид раздражителей является наиболее эффективным, к нему нервно-мышечный препарат и другие возбудимые ткани наиболее чувствительны. Таким раздражителем является электрический ток, который в связи с вышесказанным называют универсальным раздражителем.
Для наших дальнейших рассуждений о механизме возбудительного процесса очень важно обратить на это внимание. По-видимому, электрический ток в нервной, мышечной и железистой ткани запускает один и тот же процесс, который является началом и причиной возбуждения.
Для возникновения возбуждения на ткань необходимо подействовать раздражителем определенной силы, которая называется пороговой. Эта величина и является мерой возбудимости, или чувствительности, препарата к раздражителю. До достижения пороговой величины возникает местный процесс, нарастающий по мере увеличения силы раздражителя. Как только будет достигнут порог, местный процесс превращается в возбуждение, которое уже не зависит от параметров раздражителя и определяется самой возбудимой тканью.
Так, например, чтобы возбудить мышцу, достаточно подействовать на нее слабым током пороговой силы, при возбуждении же она разовьет работу, во много раз превосходящую энергию приложенного раздражителя.
Чтобы окончательно разобраться в понятиях возбудимости и порога раздражения, дадим им точное определение. Возбудимость – это чувствительность ткани к раздражению. Порог – минимальная сила раздражителя, вызывающая возбуждение. Эти две величины могут быть связаны простой формулой, демонстрирующей обратно пропорциональную зависимость между ними. Если возбудимость обозначить через Е, а пороговую силу раздражителя – через S, то Е = 1/S.
Обычно в физиологических лабораториях для измерения возбудимости применяют электрический раздражитель. При пороговых величинах он не вызывает повреждения тканей и может применяться много раз на одном и том же объекте. Кроме того, электрический ток удобно дозировать, измерять и менять форму его импульса, что во многих случаях важно для исследования.
При возбуждении в клетках физиологического объекта происходит ряд сложнейших физико-химических и биохимических процессов, с которыми мы будем знакомиться далее.
При определении минерального состава цитоплазмы клеток организма первое, что обращает на себя внимание, это – высокая концентрация в ней ионов калия и низкая – натрия. Снаружи, в омывающей клетку интерстициальной жидкости и крови, наоборот, много натрия и совсем мало калия. Что же касается кальция, магния и хлора, то этих ионов больше снаружи, чем внутри клетки.
В цитоплазме часть катионов уравновешивается органическими анионами, а не хлором. Это надо хорошо запомнить, так как в дальнейшем эти данные нам понадобятся для понимания очень важных механизмов.
«Виновником» ионной асимметрии является мембрана клетки с ее ионными каналами и насосами. Поэтому нам важно будет познакомиться с ее устройством (строением).
По современным представлениям, мембрана клетки имеет жидкостно-мозаическую конструкцию, в состав которой входят фосфолипиды, белки и углеводы. Здесь мы рассмотрим только те данные об устройстве (о строении) мембраны, которые нам понадобятся для понимания ее функции (рис. 2).
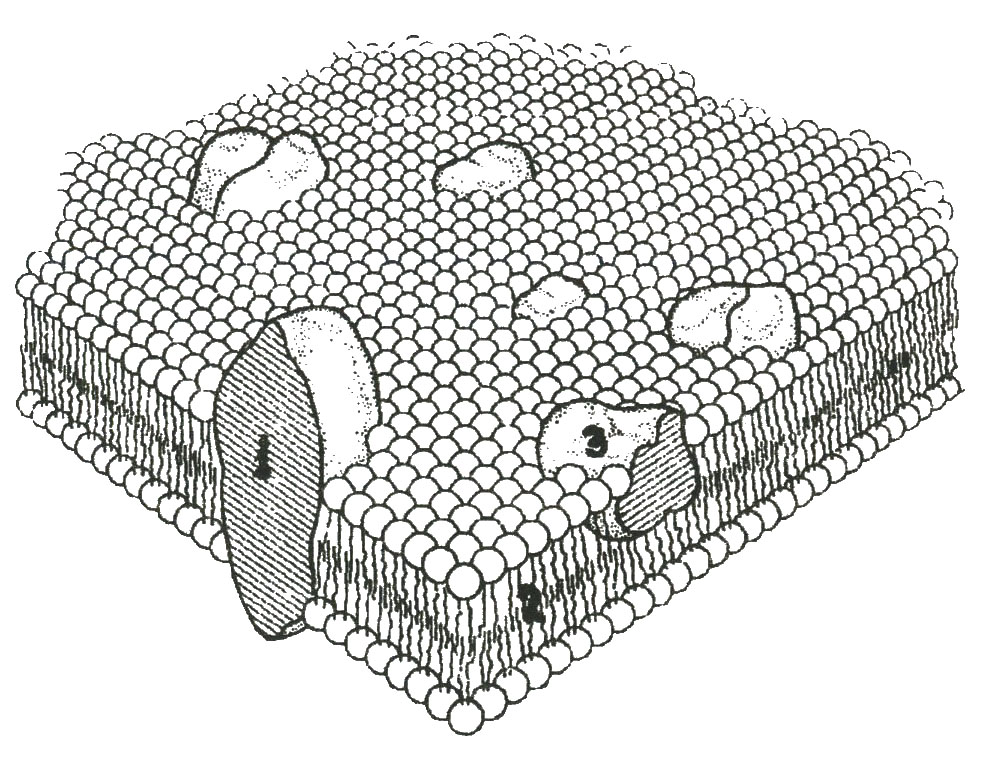
Рис. 2. Модель плазматической мембраны: 1 – интегральные белки, пронизывающие фосфолипидный бислой, 2 – фосфолипидный бислой, 3 –белки в поверхностном слое мембраны (рецепторы, ферменты)
Основой мембраны является двойной фосфолипидный слой, составляющий 30 % ее массы. Он включает в себя белки (60 %) и углеводы (10 %). Молекулы фосфолипидов построены из двух частей: гидрофильных головок и гидрофобных «хвостов». Друг с другом эти части соединяются с помощью молекулы глицерина и фосфорной кислоты. Головка фосфолипида содержит, как правило, сложное биохимическое соединение (например, спирт холин, этанол-амин, или серин), которое с помощью эфирной связи, образуемой фосфорной кислотой, соединяется с крайним углеродным атомом глицерина. Жирные кислоты, образуя эфирные связи с двумя оставшимися углеродными атомами глицерина, составляют вторую часть молекулы фосфолипида.
Попав в соответствующий водно–солевой раствор, молекулы фосфолипидов, ориентируясь «хвостами» внутрь, а головками наружу, образуют двухслойную пленку. Этот процесс называется самосборкой и часто встречается при образовании различных клеточных структур.
Соединение белковых молекул с фосфолипидным бислоем происходит довольно сложным путем. Вначале белок сорбируется на поверхности бислоя, затем происходит конформация его молекул, индуцирующая возникновение гидрофобного контакта, и, наконец, молекула белка внедряется в липидный бислой мембраны.
По характеру связи с мембраной белки подразделяются на монотопические, би- и политопические. Первые относят к периферическим белкам, вторые – к интегральным. Молекулы монотопических белков оказывают на фосфолипидный слой слабое влияние. Они довольно свободно плавают на наружной и внутренней поверхности мембраны, выполняя функцию рецепторов и ферментов. Интегральные белки пронизывают бислой насквозь и, более или менее прочно с ним связываясь, играют роль ионных каналов и ферментных систем. В зависимости от прочности связи их называют монотопическими или политопическими.
Интегральные белки существенно влияют на фосфолипидную основу мембраны, так как молекулы фосфолипидов, связываясь с белком, образуют так называемый аннулярный слой, в котором их подвижность значительно снижается. Этот процесс укрепляет бислой и фиксирует белковую молекулу.
Жирно-кислотные «хвосты» фосфолипидов обладают высокой подвижностью, особенно в тех местах, где их углеродные атомы соединяются одиночными связями. Эта подвижность снижает стабильность мембраны, и в ней появляются дефекты. Для укрепления мембраны клетка использует молекулы холестерина. Она их узнает с помощью специальных рецепторов и извлекает из находящихся в крови липопротеинов низкой плотности. Липопротеины же высокой плотности, наоборот, сорбируют холестерин из мембраны.
От содержания холестерина и насыщенности жирных кислот зависит функциональное состояние мембраны – наличие в ней дефектов и подвижность вмонтированных в нее белковых молекул. Последние могут осуществлять вращательные, поплавковые (ныряющие), линейные движения и концентрироваться в одном месте мембраны, образуя кластеры (островки). В зависимости от положения белковых молекул существенно меняется функциональное состояние клетки, ее мембранная проницаемость, чувствительность к раздражителям и ответы на действие нервных и гормональных регуляторов.
Время жизни мембранных белков составляет от двух до пяти дней, для возобновления в клетке имеется специальный механизм, синтезирующий белки и монтирующий их в соответствующие участки мембраны. Этот процесс осуществляется следующим образом. Синтез белковой молекулы начинается с сигнального пептида, способного найти на мембране рецептор, облегчающий проникновение белка в бислой. За сигнальным пептидом подтягивается рибосома, на которой синтезируется новая молекула, далее пептид отщепляется, а белок внедряется в мембрану. Рибосома отделяется, и к гидрофильной части молекулы белка присоединяются углеводные компоненты. Углеводы на мембране, как правило, соединяются с липидами (5-26%), однако наиболее важной является их кооперация с белками (10%).
В большинстве случаев молекулы углеводов локализуются на внешней поверхности мембраны, образуя своеобразные древовидные разрастания. Их функция заключается в обеспечении межклеточных взаимодействий, поддержании иммунного статуса клетки и стабилизации белковых молекул в мембране.
Мы познакомились с устройством мембраны. Чтобы разобраться в том, как она участвует в возбуждении, надо главное внимание обратить на белки, из которых построены ионные каналы. Именно эти устройства обеспечивают движение ионов через мембрану, поскольку в других местах липидного бислоя гидрофильные ионы не могут пройти.
Однако здесь следует оговориться: небольшое количество ионов проходит через дефекты мембраны и через постоянно открытые ионные каналы, но и тех, и других очень мало, в связи с чем эти потоки не имеют существенного значения.
В механизме возбуждения главную роль играют специализированные ионные каналы, обладающие особыми свойствами, позволяющими им выполнять эту важную функцию. К таким свойствам относится, прежде всего, избирательная проницаемость. Это значит, что каждый канал пропускает только свои ионы. В мембране имеются каналы, пропускающие ионы натрия, калия, кальция и хлора. Степень селективности у них различная, но свой ион всегда имеет преимущество перед чужим. Второе, не менее важное, свойство заключается в способности каналов находится в активированном и инактивированно состоянии. При активации канал открывается и пропускает ионы, при инактивации он закрывается и делается для них непроницаемым. Этот процесс осуществляется специальными заслонками, построенными из субъединиц, входящих в состав канального белка.
По типу управления заслонками все каналы можно разделить на электрические и химические. Первые открываются и закрываются под влиянием изменения заряда мембраны, вторые – при действии на нее различных химических веществ, таких, например, как медиаторы, гормоны и т.п.
По скорости активации и инактивации каналы можно разделить на быстродействующие и медленные. Последние чаще всего оказывают на электротоническое воздействие, являясь «виновниками» местных изменений ее поляризованности.
Транспорт веществ через мембрану осуществляется разными способами. Ионы могут транспортироваться независимо от других веществ, такой транспорт называется унипорт, если же они переносятся в одном направлении с другими веществами, то такой перенос называется симпорт, перенос же в противоположном направлении – антипорт.
Интересующие нас ионы транспортируются под влиянием концентрационного и электрического градиентов, то есть под влиянием электрохимического градиента. Он же создается механизмом активного транспорта за счет энергии, выделяемой при расщеплении АТФ. Такой процесс называется первично-активным транспортом, в отличие от вторично-активного, при котором перенос веществ происходит за счет энергии активно создаваемого градиента ионов – чаще всего натрия.
Нас прежде всего будет интересовать активный транспорт натрия и калия, поскольку именно от него зависит накопление калия внутри клетки, а натрия снаружи. Этот процесс осуществляется с помощью мембранного механизма, именуемого Na, K-насосом, в котором главную роль играет фермент Na,K-АТФаза.
У высших животных таким ферментом является Na, K-АТФаза типа Р, состоящая из основной субъединицы (α), которая содержит АТФ-связующий и фосфорилирующий центры, и субъединицы β, встраивающей α-субъединицу в мембрану.
Глюкозированная часть β–субъединицы располагается на наружной поверхности мембраны и, возможно, играет роль рецептора. Главная α-субъединица находится на внутренней поверхности мембраны и с помощью специального белка анкерина соединяется с цитоскелетом и митохондриями, поставляющими фермент АТФ.
Насос активируется натрием, накапливающимся внутри клетки, и калием – снаружи. Активированный фермент гидролизует АТФ, превращая ее в АДФ, а выделившаяся при этом энергия используется для переноса против концентрационного градиента натрия из клетки и калия в клетку. На каждые три иона натрия, эвакуированных из клетки, в нее поступает два иона калия, в результате цитоплазма очищается от натрия и обогащается калием.