

2 курс / Нормальная физиология / Физиология возбудимых тканей
.pdfпресинаптическую мембрану, которая отделена от субсинаптической мембраны постсинаптической области синаптической щелью. Субсинаптическая (постсинаптическая) мембрана химического синапса несколько толще, чем остальная постсинаптическая мембрана вне синаптической области. Кроме того, мембрана нервно-мышечной концевой пластинки образует регулярные субсинаптические складки, которые увеличивают площадь поверхности субсинаптической мембраны по сравнению с пресинаптической мембраной (пресинаптическим окончанием).
Рис. 31. Схема элементов нервно-мышечного синапса:
1 – миелиновая оболочка аксона; 2 – концевые веточки аксона; 3 – пузырьки, содержание ацетилхолин; 4 – митохондрия;
5 – пресинаптическая мембрана, покрывающая веточку аксона в зоне нервно-мышечного синапса; 6 – синаптическая щель;
7 – постсинаптическая мембрана, покрывающая мышечное волокно в зоне нервно-мышечного синапса; 8 – ацетилхолинорецепторы
на постсинаптической мембране; 9 – митохондрия мышечного волокна; 10 – ядро мышечной клетки; 11 – миофибрилла
В пресинаптическом окончании имеется множество синаптических пузырьков, которые содержат медиатор ацетилхолин (АХ), во время возбуждения высвобождающийся в синаптическую щель.
50
На субсинаптической мембране находятся АХ-рецепторы липопротеиновой природы, которые чувствительны к АХ и отсутствуют в соседних постсинаптических областях.
Если концевая пластинка подвергается действию АХ в течение нескольких миллисекунд, то мембрана, которая вначале деполяризуется, постепенно подвергается реполяризации, несмотря на постоянное присутствие АХ, субсинаптические рецепторы становятся рефракторными к АХ, т.е. инактивируются. Причины инактивации рецепторов пока не ясны.
4.5.2. Механизм нервно-мышечной передачи возбуждения
При достижении распространяющегося потенциала действия (нервного импульса) области пресинаптического окончания происходит деполяризация пресинаптической мембраны, которая приводит к активации электровозбудимых кальциевых каналов, их открытию и поступлению ионов кальция Са2+ внутрь окончания. Под влиянием ионов Са2+ происходит высвобождение медиатора из синаптических пузырьков, высвобождение и диффузия медиатора в синаптическую щель (несколько сотен квантов (порций) сразу). Для высвобождения каждого кванта нужны четыре иона Са2+. Если удалить Са2+, то пресинаптический потенциал действия вызовет возбуждение не сотен квантов, гораздо меньше. Квант, вероятно, содержит несколько тысяч молекул медиатора, которые выходят в очень узкую синаптическую щель в течение 1-2 мс.
Диффузия медиатора ацетилхолина к субсинаптической мембране приводит к взаимодействию его с Н-никотино-чувствитель- ными холинорецепторами субсинаптической мембраны. Присутствие Са2+ необходимо для нормального высвобождения квантов под влиянием пресинаптического потенциала действия. Степень изменения проницаемости для Са2+ зависит от величины деполяризации. Соответственно, высвобождение медиатора возрастает при увеличении амплитуды и длительности деполяризации.
Во время действия медиатора на субсинаптическую мембрану (около 1-2 мс) значительно возрастает проводимость мембраны для Na+ и К+. Мембрана мышечного волокна в области концевой пластинки (ПКП – потенциал концевое пластинки) деполяризуется до порога и возникает ПД, который распространяется по всей мышечной клетке.
51
4.5.3. Завершение действия АХ
Временно возникающий на постсинаптической мембране комплекс «АХ-рецептор» после прохождения каждого импульса разрушается ферментом ацетилхилинэстеразой. В норме действие АХ на субсинаптическую мембрану продолжается в течение короткого времени, потому что часть диффундирует из области концевой пластинки, а часть расщепляется на холин и уксусную кислоту. Продукты расщепления АХ снова поступают в пресинаптические окончания, где они участвуют в ресинтезе АХ.
4.5.4.Блокада нервно-мышечной передачи
1.Блокада проведения возбуждения в пресинаптических нервных окончаниях (например, местноанестезирующие вещества).
2.Блокада высвобождения медиатора (например, удаление из раствора Са2+ или его конкурентное замещение Мg2+или Мn2+).
3.Нарушение синтеза медиатора (например, гемихолиний, который угнетает поглощение холина пресинаптическими окончаниями).
4.Действие на субсинаптаческие АХ-рецепторы (например, кураре).
5.Угнетение холинэстеразы и, следовательно, расщепления АХ (ингибиторы холинэстеразы).
Блокада нервно-мышечной передачи используется при наркозе для исключения двигательных рефлексов и понижения мышечного тонуса.
Вещества, используемые для расслабления при наркозе или других лечебных процедурах, называются релаксантами.
4.6.Утомление нервно-мышечного препарата и утомление
вцелом организме
Утомлением называется временное понижение работоспособности клетки, органа или целого организма, наступающее в результате работы и исчезающей после отдыха.
Если длительно раздражать ритмическими электрическими стимулами изолированную мышцу, к которой подвешен не-
52

большой груз, то амплитуда ее сокращений постепенно убывает, пока не дойдет до нуля (рис. 32, а), регистрируемую при этом запись сокращений называют кривой утомления.
Рис. 32. Утомление мышцы:
А– кривая одиночного сокращения (а) и эргограмма (б) неутомленной
иутомленной мышцы; Б – локализация утомления в нервно-мышечном препарате: I – схема опыта, II – кривая утомления мышцы; а – утомление при непрямом раздражении, б – запись при прямом раздражении
Наряду с изменением амплитуды сокращений при утомлении нарастает латентный период сокращения, удлиняется период расслабления мышцы и увеличивается порог раздражения, т.е. понижается возбудимость. Все эти изменения возникают не тотчас же после начала работы мышц. При дальнейшем длительном раздражении наступает утомление мышечных волокон.
Понижение работоспособности изолированной из организма мышцы при ее длительном раздражении обусловлено двумя основными причинами. Первой из них является то, что во время сокращений в мышце накапливаются продукты обмена веществ (в частности, фосфорная кислота, связывающая ионы Са2+, молочная кислота и др.), оказывающие угнетающее влияние на работоспособность мышечных волокон. Часть этих продуктов, а также ионы калия диффундируют из волокон наружу в околоклеточное про-
53
странство и оказывают угнетающее влияние на способность возбудимой мембраны генерировать потенциалы действия.
Если изолированную мышцу, помещенную в небольшой объем жидкости Рингера, довести длительным раздражением до полного утомления, то достаточно только сменить омывающий ее раствор, чтобы восстановились сокращения мышцы.
Другой причиной развития утомления изолированной мышцы является постепенное истощение в ней энергетических запасов. При длительной работе изолированной мышцы происходит резкое уменьшение запасов гликогена, вследствие чего нарушаются процессы ресинтеза АТФ и креатинфосфата, необходимых для осуществления сокращения.
Утомление изолированной скелетной мышцы при ее прямом раздражении является лабораторным феноменом, так как в естественных условиях существования в организме возбуждающие импульсы приходят к мышце с нерва.
Нервно-мышечное соединение утомляется значительно раньше, чем мышечные волокна, в связи с чем блокирование передачи возбуждений с нерва на мышцу предохраняет ее от истощения, вызываемого длительной работой.
Доказательством того, что в системе нерв-синапс-мышца утомление развивается раньше всего в синапсе, является запись кривой утомления мышцы при непрямом ее раздражении. Как только появляются отчетливые признаки утомления, т.е. амплитуда сокращения мышцы становится заметно меньше исходной, начинают прямое раздражение мышцы и отмечают, что при прямом раздражении мышцы она начинает сокращаться с первоначальной амплитудой (см. рис. 32, б).
Нарушение нервно-мышечной передачи объясняется тем, что в нервных окончаниях при длительном раздражении резко уменьшаются запасы медиатора. Поэтому порции ацетилхолина, выделяющиеся в синапсах в ответ на каждый импульс, уменьшаются и соответственно снижаются до подпороговых величин постсинаптические потенциалы.
Наряду с этим при длительном раздражении нерва происходит под влиянием накопления в мышце продуктов обмена постепенное понижение чувствительности постсинаптической мембраны мышечного волокна к ацетилхолину. В результате
54
уменьшается величина потенциалов концевой пластинки. Когда их амплитуда падает ниже некоторого критического уровня, возникновение потенциалов действия в мышечном волокне прекращается. По этим причинам синапсы быстрее утомляются, чем нервные волокна и мышцы.
В целостном организме еще раньше нервно-мышечных соединений утомляются при работе нервные центры.
Примечание. Опыт показывает локализацию утомления в мионевральном синапсе.
Впервые И.М. Сеченов (1903) показал, что восстановление работоспособности утомленных мышц руки человека после длительной работы по подъему груза резко ускоряется, если в период отдыха производить работу другой рукой. Временное восстановление работоспособности мышц утомленной руки может быть достигнуто и при других видах двигательной активности, например при работе различных мышц нижних конечностей. В отличие от простого покоя такой отдых был назван И.М. Сеченовым активным. Эти факты доказывают, что утомление прежде всего развивается в нервных центрах.
Убедительным доказательством роли нервных центров в развитии утомления служат опыты с внушением. Так, исследуемый может длительно поднимать тяжелую гирю, если ему внушить, что в его руке находится легкая корзина. Наоборот, если внушить исследуемому, поднимающему легкую корзину, что ему дана тяжелая гиря, то быстро развивается утомление. При этом изменение пульса, дыхания и газообмена находится в соответствии не с реальной, осуществляемой человеком работой, а с той, которая ему внушена (В.М. Василевский, Д.И. Шатенштейн).
Таким образом, двигательный нервный центр, мионевральная передача являются наиболее утомляемыми в нервномышечной системе. Сама же мышца и особенно нервный проводник – звенья наименее утомляемые.
Для изучения мышечного утомления у человека в лабораторных условиях пользуются эргографами – приборами для записи амплитуды движения, ритмически выполняемого группой мышц.
55
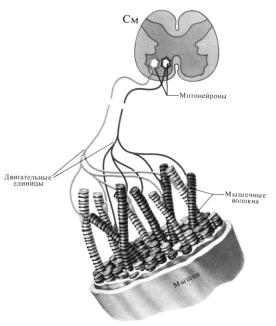
4.7. Двигательные единицы, их классификация
Анатомической и функциональной единицей скелетных мышц является двигательная единица. Под двигательной единицей следует понимать двигательный нейрон – мотонейрон, аксон и иннервируемую группу мышечных волокон (рис. 33).
Рис. 33. Строение двигательной единицы
Мышечные волокна каждой двигательной единицы (ДЕ) расположены на значительном расстоянии друг от друга. Число мышечных волокон, входящих в одну ДЕ, отличается в разных мышцах. Оно меньше в мелких мышцах, осуществляющих, тонкую и плавную регуляцию двигательной функции (например, мышцы кисти, глаза). Так, в частности, в глазных мышцах одна ДЕ содержит 13-20 мышечных волокон, а ДЕ внутренней головки икроножной мышцы – 1500-2500. Мышечные волокна одной ДЕ имеют одинаковые морфофункциональные свойства.
По морфофункциональным свойствам ДЕ делятся на три основных типа (рис. 34): I – медленные, неутомляемые, IIА –
56

быстрые, устойчивые к утомлению, IIIB – быстрые, легко утомляемые. Скелетные мышцы человека состоят из ДЕ всех трех типов. Одни из них включают преимущественно медленные ДК, другие – быстрые, третьи – и те, и другие.
Рис. 34. Двигательные единицы (ДЕ) мышцы и их типы: 1 – медленные, слабые, неутомляемые мышечные волокна.
Низкий порог активации мотонейрона; 2 – промежуточный тип ДЕ; 3 – быстрые, сильные, быстроутомляемые мышечные волокна.
Высокий порог активации мотонейрона
Медленные, неутомляемые двигательные единицы (тип I). У этих ДЕ наименьшие величины мотонейронов и соответственно наиболее низкие пороги их активности, меньшие толщина аксона и скорость проведения возбуждения по нему. Нервные волокна медленных ДЕ относятся к тонким мякотным нервным,
57
волокнам с низкой возбудимостью и малой скоростью проведения импульса.
Аксон мотонейрона разветвляется на небольшое число концевых веточек и иннервирует небольшую группу мышечных волокон. У мотонейронов медленных ДЕ низкая частота разрядов (6-10 имп/с). Они начинают функционировать уже при малых мышечных усилиях. Так, мотонейроны ДЕ камбаловидной мышцы человека при удобном стоянии работают с частотой 4 имп/с. Мотонейроны медленных ДЕ способны поддерживать постоянную частоту разрядов в течение десятков минут.
Мышечные волокна медленных ДЕ развивают небольшую силу при сокращении в связи с наличием в них меньшего, по сравнению с быстрыми волокнами, количества миофибрилл. Скорость сокращения медленных этих волокон в 1,5-2 раза меньше, чем быстрых. Основными причинами этого являются низкая активность миозин АТФазы и меньшие скорость выхода ионов кальция из саркоплазматического ретикулума и его связывания с тропонином в процессе возбуждения волокна.
Мышечные волокна медленных ДЕ развивают небольшую силу при сокращении в связи с наличием в них меньшего, по сравнению с быстрыми волокнами, количества миофибрилл. Скорость сокращения медленных этих волокон в 1,5-2 раза меньше, чем быстрых. Основными причинами этого являются низкая активность миозин АТФазы и меньшие скорость выхода ионов кальция из саркоплазматического ретикулума и его связывания с тропонином в процессе возбуждения волокна.
Мышечные волокна медленных ДЕ малоутомляемы. Они обладают хорошо развитой капиллярной сетью. Благодаря этому во время сокращения они обеспечиваются достаточным количеством кислорода. В их цитоплазме имеется большое количество митохондрий и высокая активность окислительных ферментов. Имеют большой запас гликогена и липидов. Все это позволяет выполнять работу умеренной мощности длительное время без утомления.
Быстрые, легко утомляемые ДЕ (тип II–В). Мотонейроны быстрых ДЕ крупные, имеют толстый аксон, разветвляющийся на большое число концевых веточек и иннервирующий соответственно большую группу мышечных волокон. Мотонейроны
58
обладают наиболее высоким порогом возбуждения, а их аксоны – большей скоростью возбуждения нервных импульсов.
Частота импульсации мотонейронов возрастает с ростом силы сокращения, достигая при максимальных напряжениях мышц 25-50 имп/с. Эти мотонейроны не способны в течение длительного времени поддерживать устойчивую частоту разрядов, т.е. быстро утомляются.
Мышечные волокна быстрых ДЕ, в отличие от медленных, содержат большее число сократительных элементов – миофибрилл, поэтому при сокращении развивают большую силу. Благодаря высокой активности миозиновой АТФазы у них выше скорость сокращения. Мышечные волокна быстрых ДЕ содержат больше гликолитических ферментов, меньше митохондрий и миоглобина, окружены меньшим, по сравнению с медленными ДЕ, количеством капилляров. Эти волокна быстро утомляются. Более всего они приспособлены для выполнения кратковременной, но мощной работы.
Быстрые, устойчивые к утомлению ДЕ (тип II–А). По своим морфофункциональным свойствам этот тип мышечных волокон занимает промежуточные положения между ДЕ I и II-В типов. Это сильные, быстро сокращающиеся волокна, обладающие большой аэробной выносливостью благодаря присущей им возможности использовать для получения энергии как аэробные, так и анаэробные процессы.
У разных людей соотношение числа медленных и быстрых ЕК в одной и той же мышце определено генетически и может отличаться весьма значительно. Так, например, в четырехглавой мышце бедра человека процент медленных волокон может варьировать от 40 до 98%. Чем больше в мышце процент медленных волокон, тем более она приспособлена к работе на выносливость. И наоборот, лица с высоким процентом быстрых сильных волокон в большей мере способны к работе, требующей большой силы и скорости сокращения мышц.
Таким образом, в скелетных мышцах различают быстрые и медленные двигательные единицы, состоящие соответственно из быстрых и медленных мышечных волокон.
Медленные (красные) мышечные волокна (их цвет зависит от большого содержания миоглобина) характеризуются значи-
59