

Обмен лпонп

Окисление высших жирных кислот:
Происходит путем окисления β-углеродного атома, по этому называется β-окислением. Процесс происходит в несколько этапов:
1) активация высших жирных кислот
происходит в цитозоле клетки под
действием ферментов тиокиназ с затратой
АТФ. ациладелат
ациладелат

ацилкофермент А
2) транспорт жирных кислот в митохондрии.
Сам процесс окисления жирных кислот происходит в митохондриях. Ацилкофермент А не может пройти через митохондриальную мембрану, поэтому транспорт его происходит с участием карнетина.
Карнетин – это β-гидрокси, γ,3-метиламиномасляная кислота. Под действием фермента карнитин-ацилкоферментА-ацилтрансферазы происходит перенос остатка жирной кислоты с ацилкофермента А на карнитин с образованием эфира ацилкарнитина.

ацилкарнитин
ацилкарнитин легко проходит через мембрану митохондрий. В митохондриях снова действует фермент ацилкофермент А-карнитин-ацилтрансфераза, которая переносит остаток жирной кислоты с карнитина на митохондриальный HS-KoA.
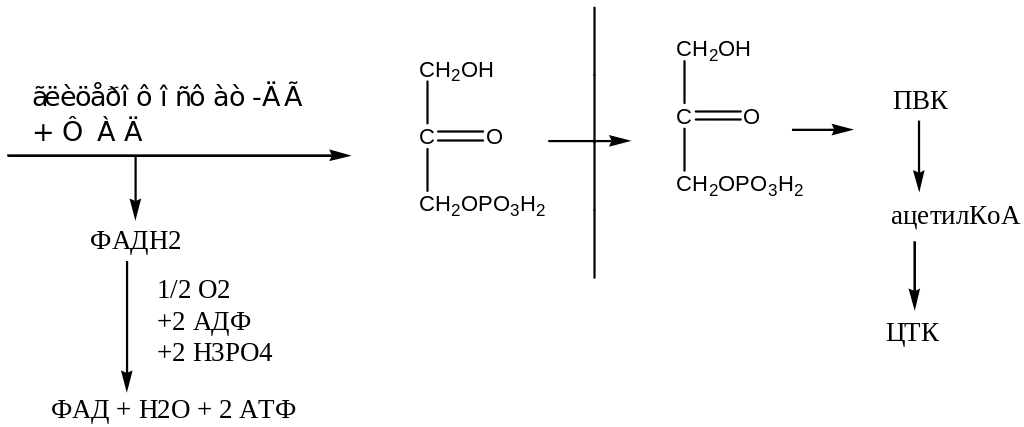
3. β-окисление жирных кислот: на ацилкофермент А действует фермент ацил-колфермент А-дегидрогеназа с простатической группой ФАД, которая восстанавливается.
O
||
R-CH2-CH2-C~SKoA ½ О2 + 2 АДФ + 2 Н3РО4
↓ ацил-КоА-ДГ + ФАД → ФАДН2 –––––––––––––––––––→ Н2О + 2 АТФ
R–C=C-C~SKoA
| | ||
H H O
Еноилацилкофермент А
|
↓ еноилацилгидротаза + Н2О
OH H O
| | ||
R-C-C-C~SKoA
| |
H H
L-β-гидроксиацил-КоА
Образуется трансизомер еноил-ацил-кофермент А. на него действует фермент еноил-ацил-гидротаза, которая присоединяет воду и образуется L-β-гидроксиацил-КоА. Этот фермент обладает стериохимической специфичностью (он действует только на трансизомеры).
На L-β-гидроксиацил-КоА действует фермент L-β-гидроксиацил-КоА-дегидрогеназа с коферментом НАД, которая обладает стериохимической специфичностью.
½ О2 + 3 АДФ + 3 Н2РО4
НАД → НАДН2 ––––––––––––––––––––→ Н2О + 3 АТФ
↓
O O
|| ||
R-C-CH2-C~SKoA
Β-кетоацил-кофермент А
|
| тиолаза + HS-KoA
↓
O O
|| ||
R-C~SKoA + CH3-C~SKoA
Ацил КоА ацетил-КоА
На β-кетоицилкофермент А действует фермент тиолаза с коферментом А, которая расщепляет его на ацил кофермент А укороченный на 2 (С) атома и ацитил кофермент А.
Ацетил кофермент А вступает в ЦТК, где окисляется до СО2 и Н2О с выделение энергии (это 4 этап).
А ацилкофермент А снова подвергается β-окислению. β-окисление жирных кислот происходит в сердце, в почках, в мышечной ткани, в легких, но наиболее активно в печени, где окисляется 60%. Только мозг не окисляет жирные кислоты для получения энергии.
Энергетический баланс жирных кислот:
Рассчитывается по уравнению: [(n/2-1)5 +12n/2]-1
Где n- число (С) атомов
n/2-1 число актов β-окисления.
5-число АТФ образовавшихся при одном акте β-окисления.
n/2 – число ацетил КоА
12 – число АТФ образующихся при окислении ацетилкофермента А в ЦТК
-1 – одна АТФ затраченная на активацию.
При окислении пальметиновой кислоты (С16) образуется 130 АТФ: [(16/2 – 1)5 +12*16/2]-1 = 130 АТФ
Синтез кетоновых тел: в печени из ацетил Ко А, который образуется при β-окислении жирных кислот, синтезируется кетоновое тело, их синтез только в печени идет.
2 ацетилкофермента А взаимодействуют с образованием ацитоацитилкофермента А.
O O ацетилкофермент А-ацетил- СH3
|| || трансфераза |
CH3-C~SKoA + CH3-C~SKoA –––––––––––––––→ C=O
↓ |
HS-KoA CH2
|
C ~ SKoA
||
O
Ацетоацетилкофермент А
O
||
CH3-C~SKoA
––––––––––––→
↓
HS-KoA
Он соединяется еще с одной молекулой ацетилкофермента А
OH O CH3
| || ОМТ-лиаза СH3 |
→COOH-CH2-C-CH2-C~SKoA ––––––––––→ | C = O
| C ~ SKoA + |
CH3 || CH2
O |
COOH
β-гидрокси, β-метил-глутарил кофермента А АУК
 АУК
АУК
НАД*Н2 - СО2
 CH3 CH3
CH3 CH3
| |
HC-OH C=O
| |
CH2 CH3
|
COOH
β-гидрокси-масляная ацетон
кислота
АУК из печени кровью доставляется к тканям (мышечная, почки, легкие, мозг). Здесь имеется фермент тиофораза (сукцинил-кофермент а-ацетоацетил-трансфераза) с помощью которого происходит взаимодействия АУК-сукцинил-кофермент А, образуется ацетоацетил кофермент А и сукцинат, который превращается в ЦТК. На ацетоацетил кофермент А действует фермент тиолаза + HS-KoA и он расщепляется на 2 молекулы ацетил кофермента А, которые окисляются в ЦТК.
CH3 COOH CH3 COOH
| | | |
C = O CH2 C = O CH2
| + | тиофораза | + |
CH2 CH2 –––––––––→ CH2 CH2
| | | |
COOH C ~ SKoA C ~ SKoA COOH
|| ↓ тиолаза + HS-KoA ЦТК
O ↓
2 СH3 – C ~ SKoA → 24 АТФ
||
O
Печень снабжает ткани хорошим энергетическим материалом, но сама печень не может использовать кетонные тела (АУК), так как не имеет фермента тиоферазы.
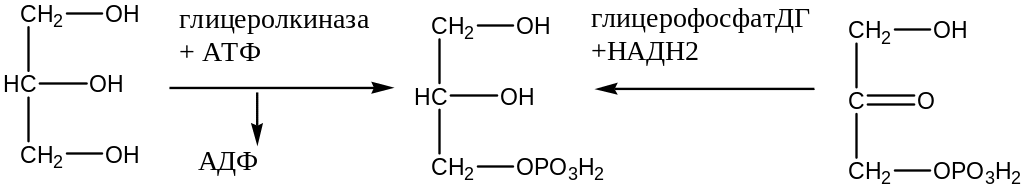
Окисление глицерина: начинается с фосфорелирования его в цитоплазме клеток за счет АТФ под действием фермента глицеролкиназы, образуется α-глицерофосфат, который сам проходит через мембрану митохондрий и в митохондриях окисляется глицерофосфат дегидрогеназой в диоксиацетонфосфат, который возвращается в цитоплазму.
Простетическая группа глицерофосфатдегидрогеназы ФАД восстанавливается и отдает протоны и электроны в полную дыхательную цепь образуется 2 АТФ.
Диоксиацетонфосфат в цитоплазме превращается по пути распада глюкозы в пируват. Пируват декарбоксилируется в ацетилкофермент А и он сгорает в ЦТК.
Энергетический баланс окисления молекуля глицерина:
2 АТФ образуются при окислении α-глицерофосфата.
2 АТФ → на анаэробных стадиях превращаются в 3 ФГА в пируват.
3 АТФ → при окислительном декарбоксилировании пирувата.
12 АТФ → ацетил кофермет А в ЦТК
3 АТФ → при окислении НАД*Н2, который образуется при окислении 3 ФГА в 1,3бисфосфоглицериновую кислоту (по малат-оксилацетальному челночному механизму)
–––––––––––––––––––––––––––––––––––
22 АТФ – 1 АТФ (затрачена на активацию) = 21 АТФ
цит МХ

α-глицерофосфат
Цит
 диоксиацетонфосфат
диоксиацетонфосфат
Синтез высших жирных кислот: ферменты этого синтеза локализованы в цитоплазме клеток в мембранах эндоплазматической сети. Для синтеза используется ацетилкофермент А, который образуется в митохондриях при окислительном декарбоксилировании пирувата и β-окислении высших жирных кислот, поэтому ацетил кофермент А должен пройти в цитоплазму, но мембрана митохондрий для него не проходима. Транспорт ацетил кофермента А в цитозоле происходит с помощью двух механизмов:
1) главный путь: ацетил кофермент А конденсируется со ЩУК под действием цитратсинтазы с образованием цитрата. Для него есть переносчик, который переносит его в цитоплазму и здесь на цитрат действует фермент цитратлиаза, которая расщепляет его на ацетил кофермент а и ЩУК.
цит
 ацетилКоА
ЩУК цитрат
ацетилКоА
ЩУК цитрат
цитрат лиаза
+ АТФ + HSKoA
––––––––––––––→ ацетилкофермент А + ЩУК
↓
АДФ + Н3РО4
2) с помощью карнитина (малопереносится).
В цитоплазме имеется фермент ацетилкофермент А- карбоксилаза, которая карбоксилирует поступивший ацетилкофермент А. это сложный регуляторный фермент, имеет кофермент биотин, активируется цитратом, ингибируется жирными кислотами (пальмитиновой кислотой.

малонилкофермент А
синтез высших жирных кислот происходит из малонилкофермента А под действием пальмитатсинтазного полиферментного комплекса, который локализован в эндоплазматической сети.
Строение пальмитатсинтазы: пальмитатсинтазный полиферментный комплекс состоит из двух одинаковых, но разнонаправленных полипептидных цепей.
Каждая полипептидная цепь состоит из 6 ферментов с разной активностью и ацил переносящего белка (АПБ). В каждой цепи есть 2 SH группы:
центральная принадлежит 4-фосфопентетину, который является простетической группой ацилпереносящего белка.
Периферическая принадлежит остатку цистеина фермента β-кето-ацилсинтазы.
Это субъединичное деление комплекса.
Кроме того деление имеется функциональное. В функционально активную частицу входит 2 фермента (β-кето-ацилсинтаза, трансацилаза) одной цепи и 4 фермента и ацилпереносящий белок второй цепи. Этим складываются центральная и периферическая SH-группы, которые работают вместе.
Механизм синтеза: фермент трансацилаза переносит остаток малонила с малонилкофермента А на центральную SH-группу и остаток ацетила с ацетилкофермента А на периферическую. Действует фермент β-кето-ацилсинтаза, которая декарбоксилирует малонил и к его остатку переносит и присоединяет ацетил с периферической SH-группой образуется β-кето-ацил-АПБ.

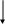
трансацилаза 2HS-KoA
β-кетоацилсинтаза
 СО2
СО2

β-кетоацилАПБ
на β-кетоацилАПБ действует кетоацилредуктаза и с помощью НАДФН2 (дает пентозный цикл распада глюкозы) восстанавливает его в β-гидроксиацилАПБ, от него фермент гидротаза отщепляет воду и образуется еноилацилАПБ, который восстанавливается еноилацилредуктазой за счет НАДФН2. образуется ацилАПБ (на SH-группе находится остаток масляной кислоты). Если в клетке нужна масляная кислота, то фермент тиоэстераза отщепляет ее от центральной группы и переносит на HS-KoA, но в основном синтез продолжается и трансацилаза переносит масляную кислоту на периферическую SH-группу, а на центральную SH-группу снова присоединяется малонил. Процесс повторяется. β-кетоацилсинтаза декарбоксилирует его и на остаток малонила переносит остаток масляной кислоты с периферической SH-группы. Образуется β-кетоацилАПБ, который содержит 6 (С) атомов.


 Кетоацилредуктаза + НАДФН2
Кетоацилредуктаза + НАДФН2
НАДФ
 β-гидрокси-ацил-АПБ
β-гидрокси-ацил-АПБ
 гидротаза
гидротаза
→ Н2О

еноилацилАПБ
еноилацилредуктаза
+ НАДФН2

бутирил-ацилАПБ
трансацилаза

↓

Процесс повторяется до тех пор пока не синтезируется пальмитиновая кислота С16.
Синтез кислот с более длинной цепью происходит путем удлинения пальмитиновой кислоты в:
ЭПС отдельными ферментами, такими же как в пальмитатсинтазном комплексе, за счет малонилкофермента А.
МХ: ферментами β-окисления. За счет ацетилкофермента А с использованием НАДФН2 для реакции восстановления.
Синтез триглицеридов (ТГ):
Происходит в жировой ткани, печени, слизистой кишечника, молочной железе.
Ферменты синтеза локализованы в эндоплазматической сети. Для синтеза используется активная форма глицерина и высших жирных кислот. Активная форма глицерина α-глицерофосфат образуется при активации глицерина за счет АТФ в печени, в почках, слизистой кишечника и диоксидацетонфосфата при его восстановлении в печени, жировой ткани, мышечной ткани.
г лицерин α-глицерофосфат диоксиацетонфосфат
печень, O печень,
почки, || жировая ткань,
слизистая кишечника +2R-C~SKoA мышечная ткань
→ 2 HS-KoA

фосфатидная кислота
фосфитаза
→ Н3РО4

диглицерид
O
||
 R-C~SKoA
R-C~SKoA
HS-KoA

триглицерин
Триглицерин человека отличается высоким содержанием олеиновой кислоты, поэтому он имеет низкую температуру плавления 16С.
Мобилизация жира из жирового депо происходит под действием импульсов из ЦНС и гормонов адреналина, глюкагона, глюкокортикоидов, инсулин тормозит липолиз и стимулирует синтез триглицеридов.
Липолиз происходит под действием трех ферментов:
триглицеридлипазы.
Диглицеридлипазы.
м
 оноглицеридлипазы
всегда в активной форме
оноглицеридлипазы
всегда в активной форме
триглицеридлипаза существует в двух формах:
а) в неактивной не фосфорелированной.
б) в активной фосфорелированной.
Фосфорелирование неактивной триглицеридлипазы происходит за счет АТФ просинкиназы, которая зависит от ц-3’-5’-АМФ. Он образуется из АТФ под действием аденилат циклазы. Ее активирует адреналин и глюкагон, поэтому они стимулируют липолиз.
Инсулин тормозит аденилатциклазу и стимулирует фосфодиэстеразу, которая расщепляет ц-3’-5’-АМФ, поэтому инсулин тормозит липолиз.