

1.4. Разнообразие и функционирование
Функционирование обычно соотносится со способностью системы осуществлять полезную работу, то есть оказывать некоторое действие на свое окружение. Для биологических систем функционирование связывается с чистой биологической продукцией. Общая продукция составляет сумму чистой продукции и затрат на основной обмен или на поддержание собственной структуры за единицу времени.
Для термодинамических систем тем больше действие, чем далее удалены они от области равновесия. Система в области равновесия обладает максимумом энтропии и минимумом действия.
Мерой способности к действию, или мерой преобразования, является H – энтропия Кульбака [Стратанович, 1985], величина которой может быть получена через сравнение равновесного рангового распределения системы с реальными. В частном случае могут быть введены локальные меры оценки отклонения реального состояния от области равновесия.
Так как в любом случае переход в ходе эволюции от нижнего иерархического уровня к верхнему приводит к росту устойчивости, определяемому более сложной структурой, то удельные затраты энергии (действий) на поддержку структуры с каждым иерархическим уровнем увеличиваются. При этом затраты на поддержку всей системы увеличиваются непропорционально увеличению ее размеров. Во всех случаях действует аллометрическая зависимость диссипации энергии
Ed = aMb ,
где М – масса;
a – диссипация энергии на единицу веса в единицу времени,
b < 1 – параметр, часто трактуемый как отношение поверхности к объему и обычно приравнивается к 0,75.
Очевидно, что это эмпирическое соотношение есть нечто иное, как другая форма записи информационного соотношения, в котором мощность сигнала определяется массой:
Cd = ln(E) = ln(a) +bln(M = b(ln(a)/b +ln(M)) = bln(a-bM).
Если положить, что шум N = ab, что точно соответствует смыслу диссипации, то получаем:
Сd = bln(M/N).
В последнем выражении не хватает в скобках только единицы, определяющей минимальный допустимый уровень мощности сигнала. Таким образом, удельная диссипация энергии есть не более как шум в степени – b , то есть в полосе частот b.
Можно полагать, что шум тем больше, чем больше информации приходится преобразовать системе для поддержки своей устойчивости. Рост структурной сложности есть логарифмическая функция от времени (t), затраченного от начала эволюции (T) до времени формирования иерархического уровня – возраста (А):
t = T-A и ln(Nt) = wln(T- A).
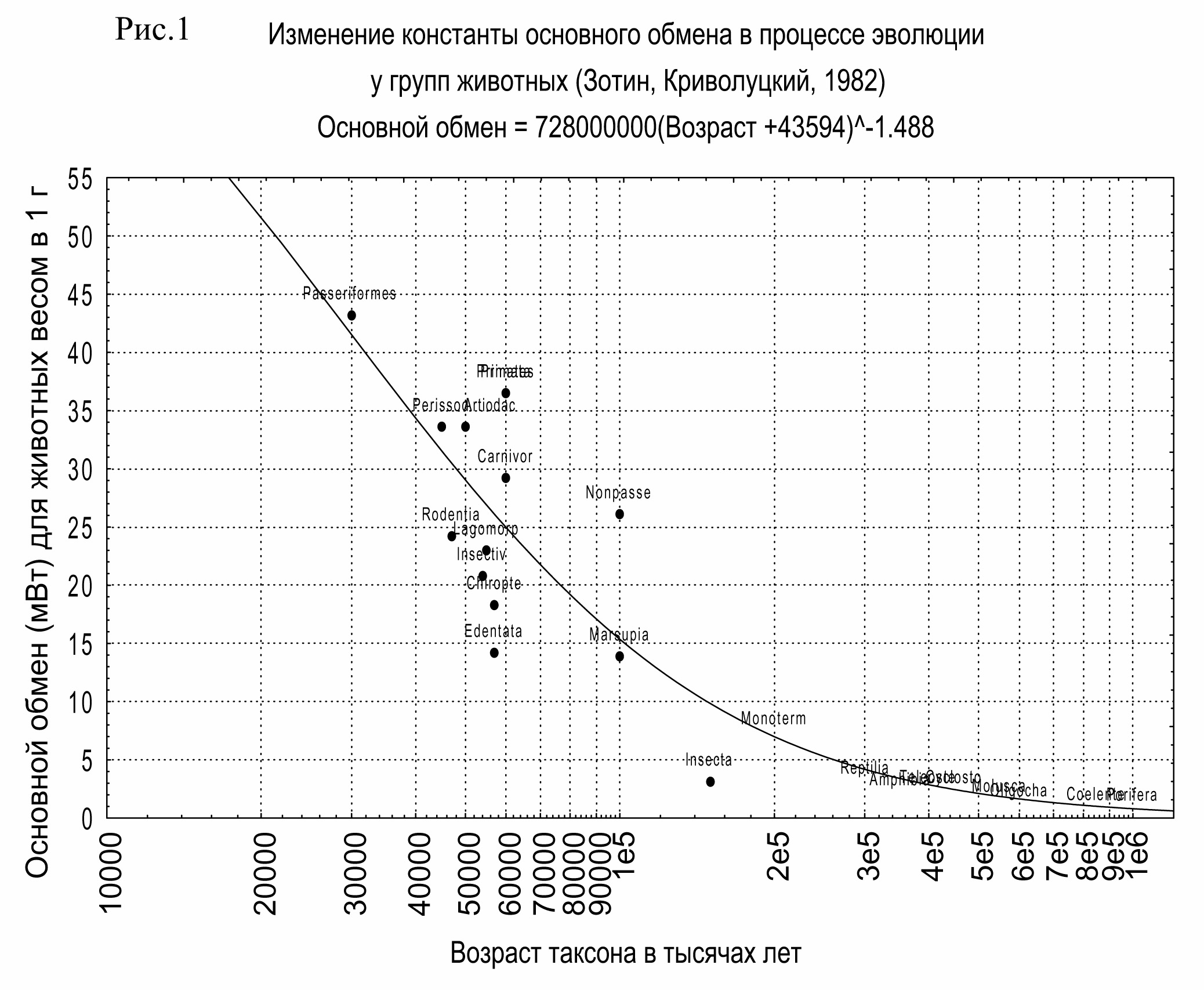
Полоса частот w в случае инвариантности отношений очевидно должна быть равна 1/b, то есть 3/2 (b = 0.75). На рис. 1 показана эмпирическая зависимость удельного основного обмена (a) как функции возраста таксона животных. Константа в показательном уравнении без учета знака равна 1,488, что неотличимо от ожидаемого значения 1,5.
Таким образом, эмпирический коэффициент точно отвечает предположениям, вытекающим из теории иерархических преобразований. Платой за повышение устойчивости является степенной рост удельной диссипации. Но при этом структурные преобразования при переходе от нижнего уровня к верхнему уменьшают затраты энергии на каждую единицу создаваемой информации. Ихкновенни на новых иерархнства ресурсов по спектру независимых частотнообразия и разнообразия лан Общий обмен с переходом от уровня к уровню растет точно по той же зависимости, что и с увеличением веса в каждой конкретной группе животных.
Внешний парадокс в соотношении: «кпд падает – устойчивость растет» разрешается тем, что создание некоторой продукции при самовоспроизводстве есть только одна и при этом не самая эффективная, а, может быть, даже наиболее примитивная, первичная форма повышения устойчивости. По мере эволюции и перехода на более высокие уровни на основе извлечения информации из взаимодействий возникают и закрепляются в памяти механизмы, обеспечивающие большую устойчивость при меньших затратах энергии.
Можно полагать, что этот общий закон эволюции разнообразия есть необходимость постоянного расширения области взаимодействий. То, что обычно определяется как «энергия»,– солнечная энергия, энергия, накопленная в форме полезных ископаемых, энергия связей атома, – есть одна из форм действий, обеспечивающая через неравновесные переходы, скачки по ступеням, усложнения организации и повышения устойчивости. По-видимому, обойти это соотношение невозможно, и потребность в энергии в ходе эволюции вместе с ростом разнообразия прогрессивно растет. В общем рост энергетических затрат определяется неизбежным увеличением в сложных структурах термодинамического первичного шума (хаоса ), не меняющего своей природы и остающимся источником новой информации.
Фактически тот же эффект проявляется и в определении связи некоторой полезной продукции (или работы) с разнообразием системы на любом уровне ее организации. Чем сложней система и выше иерархический уровень ее организации, тем больше затраты энергии на поддержание ее структуры и при прочих равных условиях ниже коэффициент ее полезного действия, или способность к производству действия (работы), но выше ее устойчивость или надежность.
Таким образом, во всех случаях, чем больше разнообразие системы или, в общем случае, ее сложность, тем больше затраты на получение от нее полезной продукции, хотя качество этой продукции может быть более высоким. Одним из важных показателей качества является устойчивость этой продукции к широкому диапазону внешних воздействий и незаменимость ее (информации) в конструкциях более высокого иерархического уровня организации.