

Електрофізіологічні явища на мембранах збудливих клітин merge
.pdfФекета Володимир Петрович |
1 |
РОЗДІЛ
1Фізіологія збудливих тканин
Збудливість – одна із фундаментальних властивостей деяких тканин живих організмів, яка полягає у їх здатності реагувати на дію зовнішніх подразників генеруванням швидких коливань заряду клітинної мембрани, здатних поширюватися на великі відстані. Завдяки цьому клітини можуть ефективно обмінюватися інформацією, інтегрувати організм в єдине ціле і пристосовуватися до змін зовнішнього та внутрішнього середовища. До збудливих тканин в організмі людини належать нервова, м’язова та частина секреторної епітеліальної тканини у залозах зовнішньої секреції. Вся нервова регуляція як на рівні центральної, так і периферичної, в т.ч. автономної нервової системи, здійснюється виключно завдяки збудливості клітин цих тканин. Вищі нервові функції (свідомість, пам’ять, навчання, інтелект) також неможливі без збудливості нейронів головного мозку.
1.Електрофізіологія мембран
Вивчення збудливості включає з’ясування таких фізіологічних феноменів як мембранний потенціал спокою, потенціал дії, локальна відповідь, рефрактерність, синаптична передача, провідність нервових волокон, збудження і скорочення скелетних та гладеньких м’язів і т.д. Розуміння цих феноменів необхідне лікарю для орієнтації в механізмах дії сучасних фармакологічних препаратів, у патогенезі ряду захворювань, у підборі та контролю ефективності немедикаментозних методів лікування і т.п. У наступних главах розділу всі ці питання розлядаються з мінімально необхідним лікарю рівнем деталізації.
1.1. Іонні канали та їх роль у процесах збудливості
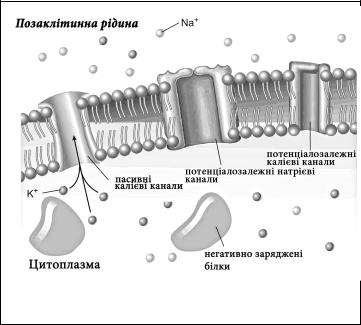
Іонні канали – це спеціальні анатомічні структури клітинної мембрани, які контролюють рух іонів через неї. Саме вони відповідальні за електричне збудження мембрани.
Фекета Володимир Петрович |
2 |
Згадаємо, що мембрана складається з біліпідного шару, у який вбудовані інтегральні та поверхневі білки. Саме інтегральні білки формують іонні канали. По суті це пори, через які можуть рухатися іони. До ключових характеристик іонних каналів відносять:
Селективність – здатність пропускати тільки певний вид іонів. Наприклад, Na+- канали пропускають тільки іони натрію, К+ – тільки іони калію, Са+2 – тільки іони кальцію і т.д. У свою чергу селективність залежить від заряду іона, його розмірів, здатності утримувати довкола себе молекули води.
Активність або пасивність – проявляється у способі відкриття каналу для руху іонів. Пасивні іонні канали відкриті постійно. До них, наприклад, відносять К+ канали, відповідальні за формування мембранного потенціалу спокою. Активні канали пропускають іони тільки при певних обставинах. Це досягається завдяки відкриттю чи закриттю воріт канала, що є фрагментом білкової молекули, здатним реагувати на дію подразників. За способом відкриття воріт активні іонні канали поділяють на хемочутливі та потенціалозалежні.
Хемочутливі канали відкривають ворота і пропускають іони при дії на них білогічно активних речовин. Такі речовини звя’зуються із рецептором мембрани і змінюють конформацію білкових молекул стінки канала. Коли вони від’єднуються від рецептора, ворота закриваються і канал припиняє пропускати іони. Прикладом біологічно активних речовин, що діють на хемочутливі
іонні канали є нейромедіатори ацетилхолін та гама-аміномасляна кислота (ГАМК). Так, ацетилхолін відкриває натрієві канали, а ГАМК – хлорні канали. Потенціалозалежні канали реагують відкриттям або закриттям воріт канала при зміні заряду їх мембрани. Так, якщо зовнішній заряд мембрани досягає - 45 mV, то відкриваються активні натрієві канали мембрани і пропускають іони натрію всередину клітини. Внаслідок цього зовнішній заряд змінюється до +30 mV, що є стимулом до закриття цих каналів і припинення транспорту натрію в клітину. Схожим чином регулюється і проникність активних потенціалозалежних калієвих каналів.
Регіональний характер розташування стосується переважної локалізації різних типів іонних каналів у мембрані нейронів. Так, пасивні канали розташовані у всіх ділянках нейрона, хемочутливі канали – у мембрані дендритів та тілі нейрона, а потенціалозалежні канали – переважно у зоні аксонного горбика, по всій мембрані немієлінізованих аксонів та у перехватах Ранв’є мієлінізованих аксонів.
Функціональна специфічність - означає спеціалізацію кожного типу іонних каналів на різних процесах, що формують збудливість. Так, пасивні іонні канали відповідають за формування мембранного потенціалу спокою, хемочутливі іонні канали забезпечують реакцію на дію зовнішніх стимулів у вигляді постсинаптичних потенціалів, а потенціалозалежні канали відповідальні за генерування та проведення вздовж мембрани потенціалу дії.

Фекета Володимир Петрович |
3 |
1.2. Мембранний потенціал спокою та механізми його формування
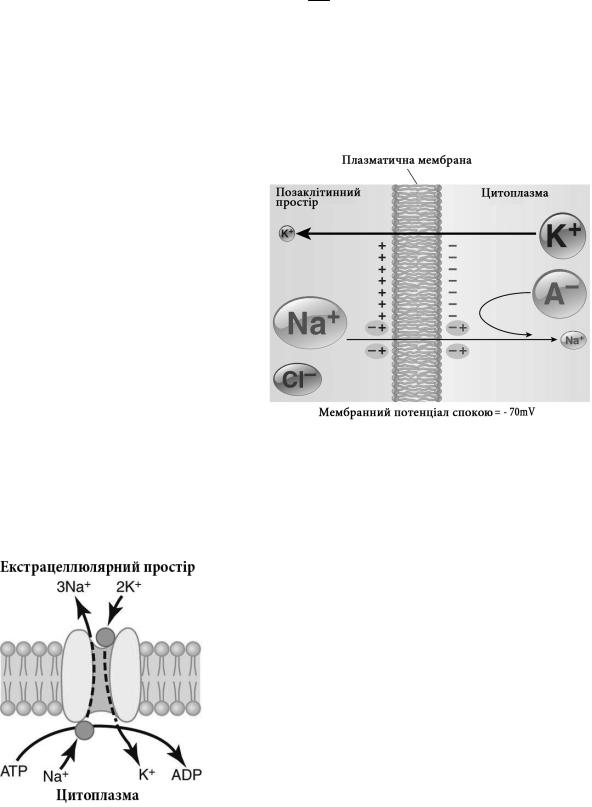
Завдяки вибірковій проникності клітинної мембрани до різних іонів та функціонуванню активних іонних насосів концентрація одних і тих же іонів у внутрішньоклітинному та позаклітинному просторі суттєво відрізняється. Цю різницю прийнято називати іонним градієнтом. У виникненні мембранного потенціалу спокою найбільше значення мають іонні градієнти калію, натрію та хлору.
Так, концентрація іонів калію у клітині приблизно у 30 разів вища, ніж у позаклітинному просторі, концентрація іонів натрію – у 10, а іонів хлору у 12,5 вища у позаклітинному просторі, ніж у клітині. В той же час, як у клітині, так і міжклітинному просторі, сумарна кількість позитивно заря-
джених іонів майже дорівнює кількості негативних. Тому в обох просторах середовище в цілому залишається електронейтральним. Основними аніонами, які балансують високий вміст калію в клітині, є негативно заряджені залишки амінокислот білків та інших органічних кислот, а надлишок катіонів калію у екстрацелюлярному просторі зрівноважується переважно аніонами хлору.
Розглянемо гіпотетичну клітину, мембрана якої проникна тільки для іонів калію. Саме такими є більшість незбудливих клітин. Концентраційний градієнт для калію є рушійною силою дифузії цих іонів із клітини на зовнішню поверхню мембрани. Однак ця дифузія одночасно створює різницю потенціалів між зовнішньою та внутрішньою сторонами мембрани, оскільки у клітині залишається більше негативно заряджених іонів, які не можуть проникнути через мембрану. Електростатична взаємодія цих іонів із катіонами калію, що вийшли із клітини перешкоджає подальшій дифузії калію.
При певному рівні різниці потенціалів між зовнішньою та внутрішньою сторонами мембрани сили дифузії зрівнюються із електричними і дифузія іонів калію припиняється.
Експериментально показано, що ця різниця потенціалів, яку називають
Фекета Володимир Петрович |
4 |
рівноважним калієвим потенціалом, складає близько - 90 mV. Знак «мінус» означає, що негативно заряджені іони превалюють на внутрішній поверхні мембрани. Математичною моделлю цього процесу є формула Нернста:
EK 61 lgCв
Сз
де ЕК – рівноважний калієвий потенціал; Св – внутрішньоклітинна концентрація іонів К; Сз – зовнішньоклітинна концентрація іонів К.
У збудливих клітинах, зокрема у нейронах, ситуація дещо складніша у зв’язку з тим, що їх мембрана в стані спокою проникна не тільки для іонів калію, але в меншій мірі
– для іонів натрію завдяки пасивним |
|
|
іонним каналам. Сукупність хіміч- |
|
|
|
||
них та електричних сил, що зумов- |
|
|
люють рух іонів через мембрану, на- |
|
|
зивають |
електрохімічним градієн- |
|
том. Якщо електрохімічний градієнт |
|
|
для калію спричинює його вихід на |
|
|
зовнішню поверхню мембрани, то |
|
|
електрохімічний градієнт для натрію |
|
|
навпаки призводить до його транс- |
|
|
порту в клітину. Цьому сприяє як бі- |
|
|
льша концентрація натрію у позаклі- |
|
|
тинному просторі порівняно із внут- |
|
|
рішньоклітинним, так і електроста- |
|
|
тичне притягання позитивно заря- |
|
|
джених |
іонів натрію негативними |
|
аніонами, що локалізовані на внут- |
|
|
рішній стороні клітинної мембрани. |
Мал.1.4. Механізм формування мембранного |
|
Потрібно відмітити, що кількість на- |
потенціалу спокою |
|
трієвих каналів на мембрані значно |
|
|
|
||
менша, ніж калієвих, тому зустрічний потік катіонів натрію в клітину незначний в порівнянні з виходом калію із клітини.
Тим не менше, цей потік зменшує трансмембранну різницю потенціалів до приблизно – 70
|
mV. Оскільки ця різниця менша за рівноважний ка- |
|
лієвий потенціал, то дифузійний градієнт для калію |
|
|
|
перевищує сили електростатичної взаємодії і спри- |
|
чинює постійний вихід цього іона на зовнішню сто- |
|
рону клітинної мембрани. Для компенсації втраче- |
|
них відповідно внутрішньота зовнішньоклітинним |
|
середовищем іонів калію і натрію необхідним є по- |
|
стійне функціонування Na-K помпи – специфічного |
|
інтегрального білка клітинної мембрани, який за ра- |
|
хунок гідролізу АТФ транспортує 3 іони Na+ із клі- |
|
тини в обмін на транспорт 2-х іонів К+ у клітину. |
|
Описана вище динамічна рівновага між різнонапра- |
|
вленим рухом іонів натрію та калію в результаті фо- |
|
рмує характерну для кожної збудливої клітини різ- |
Мал.1.5. Натрієво-калієва |
ницю потенціалів між зовнішньою та внутрішньою |
сторонами мембрани, яку позначають терміном |
|
помпа клітинної мембрани |
мембранний потенціал спокою (МПС). У деяких |
|
збудливих клітинах, як, наприклад, у міоцитах гла- |
|
Фекета Володимир Петрович |
5 |
деньких м’язів, поряд із іонами натрію і калію у виникненні МПС відіграють аніони хлору, а його рівень в збудливих клітинах різних тканин може знаходитися в межах від -30 до
-90 mV.
1.3. Потенціал дії та його фази
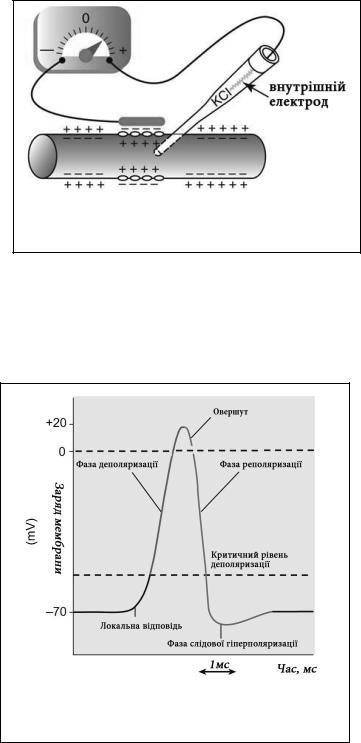
Потенціалом дії (ПД) називають швидкі зміни заряду мембрани, що виникають під дією порогових стимулів, і здатні розповсюджуватися вздовж мембрани збудливих клітин. Завдяки ПД збудливі клітини можуть обмінюватися інформацією між собою та іншими клітинами організму, що в свою чергу забезпечує інтеграцію організму в єдине ціле та його пристосування до зовнішнього середовища. Механізм формування ПД пов’язаний із активними іонними натрієвими та калієвими каналами, які за долі секунди змінюють проникність клітинної мембрани збудливих клітин для цих іонів. В експерименті ПД можна зареєструвати на осцилографі з допомогою двох електродів.
Один з нихактивний розташовують у внутрішньоклітинному середовищі, а інший – пасивний прикладають до зовнішньої сторони клітинної мембрани.
Розглянемо більш детально процеси, які відбуваються при формуванні ПД на прикладі збудження типового еферентного нейрону. Початковим місцем виникнення збудження є аксональний горбик – особлива ділянка у місці відходження аксона від тіла нейрона, яка найбільш чутлива до зовнішніх стимулів завдяки високій порівняно з іншими ділянками мембрани щільності активних натрієвих каналів.
Зміни заряду мембрани аксонного горбика при дії подразників формують 3 послідовні фази ПД:
Фаза деполяризації виникає |
|
||
у тому разі, якщо подразник змінив |
|
||
рівень заряду мембрани від рівня |
|
||
мембранного потенціалу |
спокою |
|
|
до так званого критичного рівня |
|
||
деполяризації, який зазвичай на 15- |
|
||
30 mV ближчий до нульового заря- |
|
||
ду мембрани, ніж МПС. В більшо- |
|
||
сті нейронів він близький до – 45 |
|
||
mV. Такий зсув заряду мембрани |
|
||
пов’язаний із відкриттям активних |
|
||
потенціалозалежних натрієвих ка- |
Мал.1.7. Типовий потенціал дії нервової кліти- |
||
налів і дифузією іонів натрію все- |
|||
ни та його фази |
|||
редину клітини. Оскільки |
ці іони |
||
|
|||
Фекета Володимир Петрович
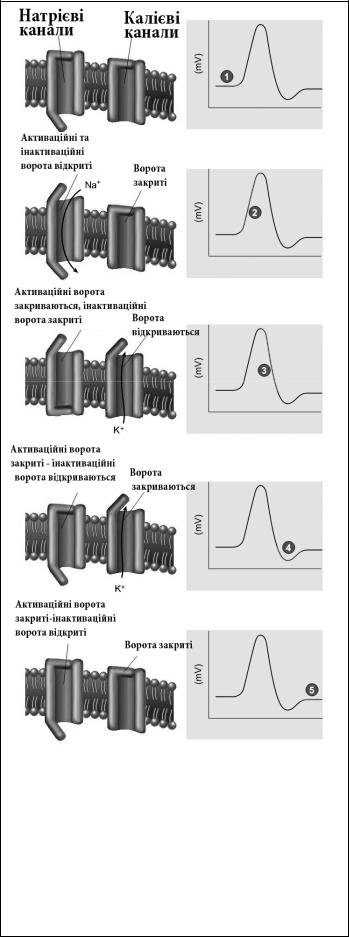
Мал.1.8. Динаміка стану воріт натрієвих та калієвих каналів в процесі генерування потенціалу дії. Цифрами позначено:
1- мембранний потенціал спокою;
2- фаза деполяризації;
3- фаза реполяризації;
4- фаза слідової гіперполяризації;
5- повернення заряду мембрани до рівня мембранного потенціалу спокою
6
мають позитивний заряд, то вони зменшують різницю потенціалів між зовнішньою та внутрішньою сторонами мембрани, що створює позитивний зворотний зв’язок із сусідніми незбудженими каналами. Цей процес відносно повільний до моменту досягнення критичного рівня деполяризації, а після цього розгортається лавиноподібно аж до моменту досягнення мембраною рівня заряду +30 mV. Тривалість фази деполяризації у нейронах знаходиться у межах 0,2-1,0 мс. Проте у інших збудливих клітинах, зокрема скелетних та гладеньких м’язах і міокарді вона може тривати у десятки разів довше. Ділянка ПД, що розташована на його графіку вище нульової лінії, називається овершутом. ПД у деяких збудливих клітинах, зокрема секреторних клітинах екзокринних залоз та міоцитах гладеньких м’язів, може бути позбавлений овершуту.
Фаза реполяризації розпочинаєть-
ся відразу після досягнення пікового рівня заряду внутрішньої мембрани і продовжується до моменту повернення цього заряду до рівня МПС. Молекулярні механізми цієї фази пов’язані із інактивацією потенціалозалежних натрієвих каналів та одночасним відкриттям потенціалозалежних калієвих каналів мембрани. Встановлено, що у натрієвих каналах існують два види воріт: на зовніщній стороні мембрани – активаційні ворота, а на внутрішній – інактиваційні ворота. Саме закриття останніх воріт завершує фазу деполяризації і поступово припиняє потік іонів натрію в клітину. Протягом фази реполяризації активаціні ворота поступово відновлюють свій початковий стан і в кінці цієї фази знову перебувають у закритому стані. Одночасно з цим відбувається дифузія катіонів калію через відкриті активні калієві канали. Вихід позитивно заряджених іонів на зовнішню поверхню мембрани відновлює її початковий рівень поляризації. Однак цей процес триває дещо довше, ніж фаза деполяризації, що пояснюється дещо меншою щільністю цих каналів та їх вищою інертністю. В середньому у більшості нейронів фаза реполяризації триває 1-2 мс.
Фекета Володимир Петрович |
7 |
Фаза слідової гіперполяризації формується протягом наступних 2-3 мс після завершення фази реполяризації і пов’язана із додатковим виходом іонів калію на зовнішню поверхню мембрани через частину ще не повністю закритих активних калієвих каналів. Протягом цієї фази завершується відновлення початкового стану активаційних та інактиваційних воріт активних натрієвих каналів і закриття воріт усіх потенціалозалежних калієвих каналів. Заряд мембрани знову повертається до рівня МПС.
Відповідність стану іонних каналів мембрани та поведінки їх воріт фазам ПД детально проілюстрована на мал. 1.8.
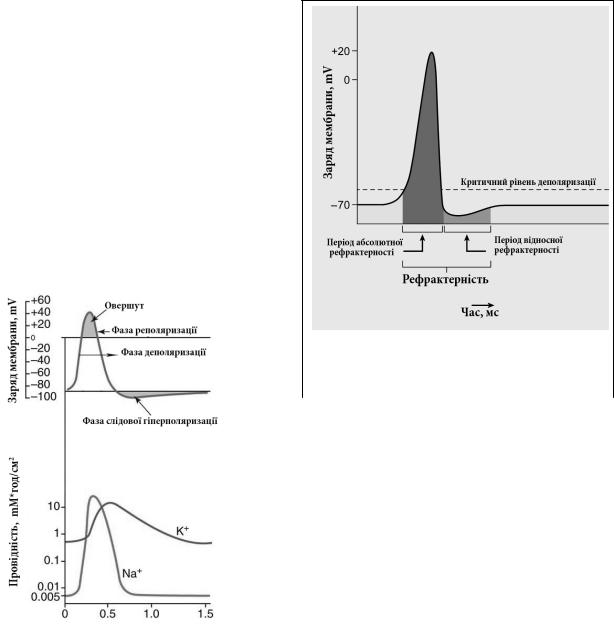
1.4. Рефрактерність збудливих клітин та її механізми
Експериментально встановлено, що при багатократній зовнішній стимуляції збудливої клітини пороговими подразниками, тобто такими – що здатні деполяризувати клітинну мембрану до критичного рівня, вона може повторно збуджуватися і генерувати ПД тільки до певної частоти стимуляції. Якщо ж цю частоту підвищувати, то клітина деякий час реагує утворенням ПД тільки у відповідь на надпорогові стимули, а при подальшому збільшенні частоти стимуляції вона не реагує на подразники будь-якої сили.
Нездатність клітинної мембрани відповідати на порогове подразнення ге-
|
|
Мал.1.9. Співвідношення фаз потенціалу |
|
|
|
дії із періодами рефрактерності клітинної |
|
|
|
мембрани |
|
|
|
|
|
|
неруванням ПД називають рефрактерністю. Ре- |
||
|
фрактерність поділяють на абсолютну – мембрана |
||
|
не реагує на подразник будь-якої сили та відносну |
||
|
– ПД виникає тільки при дії на мембрану надпоро- |
||
|
гових подразників. За тривалістю абсолютна ре- |
||
|
фрактерність у нейронах приблизно відповідає су- |
||
|
мі фаз деполяризації та реполяризації, а відносна – |
||
|
фазі слідової гіперполяризації. |
||
|
|
Молекулярні механізми рефрактерності |
|
|
пов’язані із станом активаційних воріт потенціало- |
||
|
залежних натрієвих каналів. Так, протягом фази |
||
Мал.1.10. Співвідношення фаз |
деполяризації ворота пропускають у клітину лави- |
||
ноподібний потік іонів натрію, а після початку фа- |
|||
потенціалу дії із провідністю |
|||
зи реполяризації поступово відновлюють свій по- |
|||
мембрани для іонів натрію та ка- |
|||
чатковий стан, закриваючи вхід у канал для іонів |
|||
лію |
|||
|
|
||

Фекета Володимир Петрович |
8 |
натрію, а в кінці фази реполяризації знову набувають здатності реагувати на дію подразників. Однак необхідно врахувати, що при дії подразника у фазі слідової гіперполяризації вхід позитивно заряджених іонів натрію конкурує із вихідним потоком катіонів калію через відносно повільні потенціалозалежні калієві канали, які відновлюють свій початковий стан тільки в кінці цієї фази. Тому для досягнення критичного рівня деполяризації і генерації ПД сила подразника повинна біти більшою, ніж звичайно. Співвідношення між фазами ПД та періодами рефрактерності проілюстровано на мал. 1.10.
В клініці рефрактерність збудливих структур (периферичних нервів, синапсів, скелетних м’язів) оцінюють шляхом визначення їх лабільності, мірою якої є максимальна частота нав’язаної зовнішньої стимуляції, яку ця структура здатна відтворити у вигляді послідовності ПД. Якщо, наприклад, певна структура на частоту стимуляції 200 імпульсів за секунду відповідає генеруванням 200 ПД за секунду, то її лабільність складає 200 імп/c. Зрозуміло, що чим коротший ПД та період абсолютної рефрактерності, тим вищою є лабільність збудливої структури. Тому, зазвичай, найбільшу лабільність мають нерви, дещо меншу – нейрони, ще меншу – синапси, і порівняно низьку – скелетні та гладенькі м’язи.
Абсолютна рефрактерність має винятково важливе значення для якості передачі інформації між збудливими клітинами. Оскільки дискретним носієм цієї інформаціїї є ПД, то його передчасне завершення або деформація неминуче впливали б на точність нервого коду, що, як відомо визначається кількістю ПД в пакеті нервових імпульсів та їх частотою. Період абсолютної рефрактерності гарантує завершення двох ключових фаз ПД – деполяризації та реполяризації, а відносна рефрактерність надає необхідної гнучкості у кодуванні надпорогових стимулів.
1.5. Локальна відповідь та її відмінності від потенціалу дії
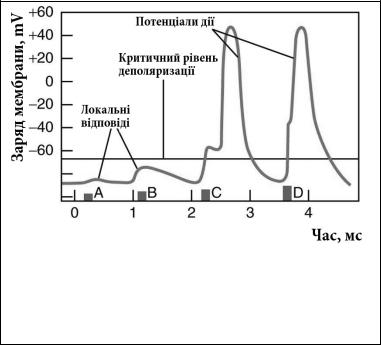
Виникає питання: як буде вести себе мембрана збудливих клітин при її стимуляції підпороговими подразниками? Експериментально встановлено, що на дію таких подразників мембрана реагує зсувом заряду в сторону деполяризації або ж гіперполяризації, але не генерує при цьому повноцінний ПД. Такі реакції мембрани називають локальною відповіддю (ЛП). ЛП можна отримати не тільки в експерименті. В інтактному організмі вони забезпечують синаптичну передачу, сприйняття інформації рецепторами, гальмування в центральній нервовій системі і т.д. Тому важливо розуміти їх властивості та механізми.
Локальна відповідь у трьох аспектах суттєво відрізняється від потенціалу дії. Поперше, вона підлягає закону сили – у певних межах амплітуда ЛВ прямо пропорційно залежить від сили подразника. В той же час ПД підпорядковується закону «все або нічого» – у відповідь на дію порогових і надпорогових стимулів виникає ПД максимально можливої амплітуди, а на дію підпорогових – не виникає взагалі. По-друге, ЛП не поширюється
Мал.1.11. Зміни заряду мембрани при локальній відповіді а) локальна деполяризація; б) локальна гіперполяризація
Фекета Володимир Петрович |
9 |
вздовж мембрани збудливих клітин, що й дало підстави називати цю відповідь локальною , а ПД – без затухання поширюється на далекі відстані. По-третє, ЛП на відміну від ПД, здатна до сумації, яка буває часова та просторова. Часова сумація спостерігається при умові короткого проміжку часу між послідовними стимулами, а просторова – при одночасному надходженні до обмеженої ділянки мембрани декількох підпорогових подразників. Із цим явищем ми познайомимося детальніше при розгляді механізмів синаптичної передачі.
Іонні механізми ЛП пов’язані із функціонуванням активних натрієвих та каліє-
вих каналів мембрани. Ці канали, як уже згадувалося раніше, можуть бути або хемочутливими, або потенціалозалежними. Локальна відповідь в сторону деполяризації, як правило, спостерігається при відкритті під дією підпорогових стимулів воріт натрієвих каналів мембрани. Рух катіонів натрію у цитоплазму зменшує різницю потенціалів між зовнішньою та внутрішньою сторонами мембрани, але не в такій мірі, яка б забезпечила досягнення критичного рівня деполяризації, необхідного для розвитку ПД. Локальна відповідь в сторону гіперполяризації зазвичай виникає при відкритті воріт активних калієвих каналів. Вихід калію із цитоплазми на зовнішню поверхню мембрани призводить до суттєвого зростання мембранного потенціалу спокою. Така мембрана має значно нижчу збудливість, ніж при нормальному значенні МПС, оскільки для її деполяризації до критичного рівня необхідний надпороговий подразник. У синапсах ЛП можуть бути пов’язані з рухом не тільки натрію й калію, а й інших іонів, зокрема – кальцію та хлору.
1.6. Проведення потенціалів дії вздовж мембрани збудливих клітин
Однією із найважливіших властивостей ПД є його здатність бездекрементно (тобто без затухання) проводитися вздовж мембрани збудливих клітин. На проведенні ПД спеціалізуються відростки нервових клітин – дендрити та аксони. Але мембрана соми також здатна проводити збудження.
Для прикладу розглянемо механім проведення ПД у немієлінізованих нервових волокнах. Такими, зокрема, є аксони більшості постгангліонарних нейронів автономної нервової системи (мал.1.13). Якщо нанести порогове подразнення у певному місці такого волокна, то збуджена ділянка протягом фази деполяризації буде мати протилежний заряд по відношенню до сусідньої незбудженої ділянки. При цьому між цими ділянками виникне різниця потенціалів, яка спричинить рух іонів в напрямку до протилежно зарядженої ділянки. Цей рух іонів прийнято називати локальним електричним струмом, який виконує роль порогового подразника для сусідньої незбудженої ділянки. Такі струми розповсюджуються в обидві сторони від місця первинного подразнення, тому кажуть, що нерви
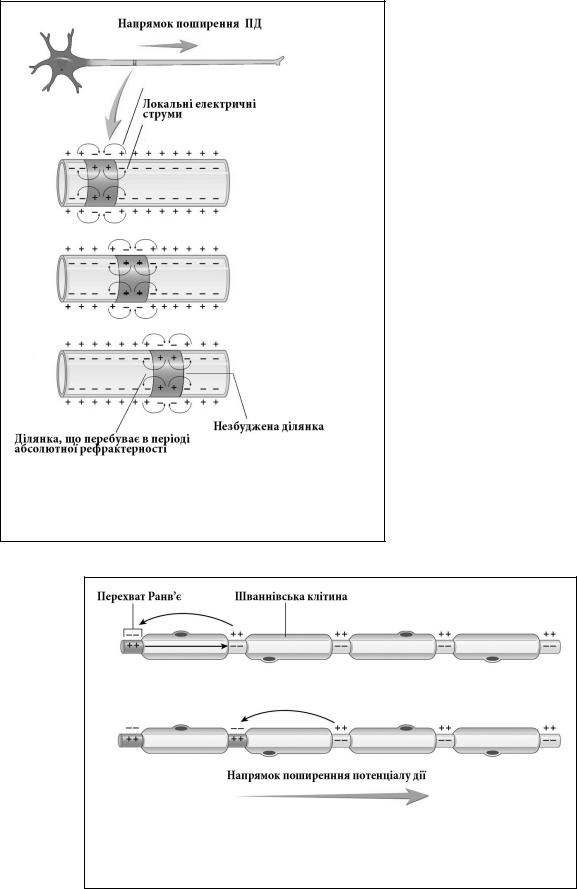
Фекета Володимир Петрович |
10 |
мають двохсторонню провідність. Сусідні ділянки в свою чергу деполяризуються, генерують ПД і передають «естафету» далі. Такий спосіб проведення збудження називають безперервним. Він є відносно повільним і енергозатратним, оскільки на підтримання постійного рівня МПС після чергового ПД потрібна енергія, яка витрачається переважно на функці-
онування Na-K помпи.
Дещо інший механізм проведення ПД має місце у мієлінізованих волокнах соматичної та автономної нервової системи (мал.1.14). Специфіка цих волокон полягає в тому, що більша частина аксонів у їх складі щільно прикрита спірально закрученими Шваннівськими клітинами. Ці клітини містять жироподібну речовину – мієлін, яка є хорошим ізолятором і запобігає руху будьяких іонів через мембрану. Можна вважати, що у місці прилягання мієлінової оболонки мембрана аксона не збуджується взагалі. Однак між окремими Шваннівськими клітинами є вільні від мієліну проміжки – перехвати Ранв’є, які до того ж мають високу в порівнянні з безмієліновими волокнами концентрацію потенціалозалежних натрієвих каналів. Спосіб проведення ПД у таких волокнах нагадує проведення ПД безмієліновими волокнами з тією різницею, що локальні електричні струми виникають не між су-
Мал.1.14. Сальтаторний механізм проведення збуджень мієлінових нервових волокнах