

Биофизика 02
.pdfлінійну залежність електричного потенціалу ϕ від координати х (так зване наближення постійного струму), тобто
де ∆ϕ — різниця потенціалів по обидва боки мембрани, l — товщина мембрани. Тоді:
Після розділення змінних одержуємо:
Проінтегруємо це рівняння, використовуючи граничні умови: при
х = 0 с =  , при х = l с =
, при х = l с =  :
:
або
звідки одержуємо вираз для потоку:
Концентрації іонів усередині мембрани пропорційні відповідним концентраціям у водяних розчинах (див. 5.3), тобто
 = Kco;
= Kco;  = Kci.
= Kci.
Якщо врахувати, що u = D/RT і P = DK/l, то рівняння для пасивного потоку іонів перепишеться у вигляді (рівняння Гольдмана):
(5.6) Рівняння Гольдмана (5.6) дозволяє обчислити величину пасивного потоку іонів, якщо відомі концентрації іонів з обох боків мембрани, різниця потенціалів і коефіцієнт проникності, який, як правило, виз-
начають в експерименті методом мічених атомів.
Потік J у рівнянні Гольдмана (5.6) є сумою двох потоків, один із яких направлений із клітини назовні (Jo), а другий із міжклітинного простору в клітину (Ji). Так як ці потоки мають протилежний напрямок, то один з них береться зі знаком «мінус»:
101

J = Jo – Ji.
Перепишемо рівняння (5.6) у вигляді:
Розділивши сумарний потік у рівнянні (5.6) на односторонні складові, одержимо вирази для Jo і Ji:
Так як потік, направлений усередину клітини, залежить лише від позаклітинної концентрації іонів, а потік, направлений назовні, — від внутрішньоклітинної, то ці рівняння доводять незалежність вхідного і вихідних потоків у випадку пасивного транспорту за умови лінійної зміни електричного потенціалу уздовж товщини мембрани.
Відношення протилежно направлених потоків описує рівняння Уссинга — Теорелла:
(5.7) Пасивний транспорт може відбуватися лише у випадку наявності на мембрані градієнтів концентрації або електричного потенціалу (тут мається на увазі, що перенесення речовини не відбувається, коли сумарний потік дорівнює нулю: J =Jo – Ji =0, хоча окремі його компоненти, як правило, відмінні від нуля) і описується в загальному випадку рівняннями Нернста — Планка (5.5), Гольдмана (5.6) або Уссинга — Теорелла (5.7), невиконання яких свідчить про те, що перенесення іонів здійснюється активно або через вузькі мембранні канали, де неможливе існування
незалежних іонних потоків.
§25. Рівняння Нернста та рівновага Доннана
Уживому організмі по обидва боки клітинної мембрани (у клітині та в позаклітинному середовищі) знаходяться іонні розчини, причому концентрації того самого іона по обидва боки мембрани часто відрізняються. Однією з причин встановлення і підтримки цих градієнтів концентрацій є різна проникність мембрани для тих чи інших іонів. У цьому випадку градієнт концентрації однієї речовини може підтримуватися за рахунок наявності градієнту концентрації іншої речовини і/або різниці потенціалів по обидва боки мембрани (так званої трансмемб-
102

ранної різниці потенціалів).
Умовою встановлення рівноваги між двома розчинами, розділеними полупроникною мембраною (проникною для молекул розчинника і непроникною для іонів), із різними концентраціями однакових іонів є рівність електрохімічних потенціалів по один та другий бік мембрани:
де ci(co) і ϕi(ϕo) — відповідно концентрації іона та електричні потенціали усередині (зовні) клітини. Тут µ0i = µ0o, так як ми вважаємо, що з іоном по обидва боки мембрани не відбувається ніяких хімічних перетворень, і він знаходиться в тому самому розчиннику.
Звідси можна одержати значення різниці електричних потенціалів, яка встановлюється на мембрані при наявності градієнту концентрацій
(рівняння Нернста):
(5.8) Як бачимо, причиною різниці електричних потенціалів на мембрані
є градієнт концентрацій. Якщо ci = co, то ∆ϕ = 0.
Розглянемо випадок, коли по обидва боки мембрани знаходяться іони калію і хлору різних концентрацій. Рівноважний мембранний потенціал для іонів калію має вигляд:
(5.9)
а для хлору —
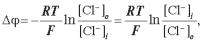 (5.10) де [K+]i і [Cl–]i ([K+]o і [Cl–]o) — концентрації відповідних іонів усередині (зовні) клітини. Дорівняємо рівняння (5.9) і (5.10):
(5.10) де [K+]i і [Cl–]i ([K+]o і [Cl–]o) — концентрації відповідних іонів усередині (зовні) клітини. Дорівняємо рівняння (5.9) і (5.10):
(5.11)
де r — відношення Доннана.
З виразів (5.8) і (5.11) видно, що градієнт концентрацій по обидва боки мембрани може підтримуватися наявністю трансмембранної різниці потенціалів, або, навпаки, різниця потенціалів визначається наявністю градієнту концентрацій, крім того, трансмембранна різниця концентрацій однієї речовини може підтримувати різницю концентрацій іншої речовини. Наприклад, в еритроцитах наявність градієнтів концентрацій іонів створює трансмембранний потенціал; у м’язових клітинах Na+,K+Fнасоси за рахунок енергії АТФ створюють градієнт
103
концентрацій іонів калію, що викликає, відповідно, появу трансмембранного потенціалу, а це, у свою чергу, підтримує градієнт концентрації іонів хлору.
У клітині крім малих іонів знаходяться ще і заряджені макромолекули (білки, нуклеїнові кислоти). Розглянемо випадок, коли в системі знаходяться іони калію і хлору і негативно заряджені макромолекули, причому мембрана проникна для малих іонів і непроникна для великих молекул. Для кожного з розчинів, що знаходяться по обидва боки мембрани, необхідне виконання умови електронейтральності, тобто сума позитивних і негативних зарядів всіх іонів повинна дорівнювати нулю. Отже,
|
|
|
|
[K+] |
= [Cl–] |
+ [R–] |
|
, |
(5.12) |
||
|
|
|
|
|
i |
|
i |
i |
|
|
|
|
|
|
|
[K+] |
o |
= [Cl–] |
o |
+ [R–] |
o |
, |
|
|
|
|
|
|
|
|
|
|
|||
де [R–] |
i |
і [R–] |
о |
— концентрація макромолекул у внутрішньоF і позаклі- |
|||||||
тинному середовищі. Міжклітинна рідина майже не містить великих
заряджених частинок ([R–] |
о |
≈ 0), тому |
|
|
|
||
|
|
[K+] |
o |
= [Cl–] |
o |
= с. |
(5.13) |
Поєднавши (5.13) та (5.11), одержимо:
[K+]i [Cl–]i = [K+]o [Cl–]o = с2.
Підставимо сюди умову електронейтральності для внутрішньоклітинного середовища (5.12):
{[Cl–]i + [R–]i}[Cl–]i = с2.
Розв’язавши це квадратне рівняння, знайдемо, що концентрація малих аніонів у внутрішньоклітинному середовищі складає:
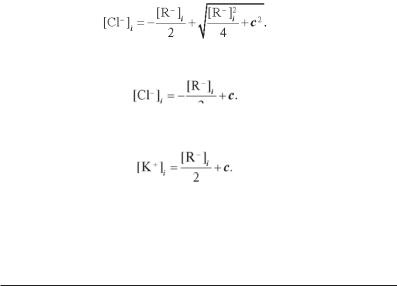
Якщо концентрація макромолекул значно нижче концентрації малих іонів ([R–]i << с), то в цьому випадку одержуємо:
 (5.14) Тоді з виразів (5.12) і (5.14) випливає, що внутрішньоклітинна кон-
(5.14) Тоді з виразів (5.12) і (5.14) випливає, що внутрішньоклітинна кон-
центрація іонів калію складає:
Тобто для компенсації негативних зарядів макромолекул частина малих негативних іонів виходить із клітини (у даному випадку зменшується концентрація іонів хлору), а частина позитивних зарядів проходить у клітину з міжклітинного середовища (збільшується концентрація іонів калію).
104

Виникаюча на мембрані доннанівська різниця потенціалів складає:
Так як ln(1 – x) ≈ –x, то
Як правило, заряд макромолекули дорівнює не одиниці, як було зазначено вище, а 10–20, а іноді і значно більше. Позначивши цю величину n, можна показати, що трансмембранна різниця потенціалів буде дорівнювати
Наприклад, якщо концентрація макромолекул складає 0,01с, n =10, то доннанівська різниця потенціалів при температурі 37 °С складе —1,3 мВ.
§ 26. Іонний транспорт через канали
Транспорт іонів через ліпідний бішар за допомогою простої дифузії відбувається вкрай повільно. Проникнення одного моля іонів із розчину з діелектричною проникністю εв ≈ 80 у мембрану з діелектричною проникністю εм ≈ 2 супроводжується зміною їхньої вільної енергії на величину (формула Борна):
(5.15) де z— заряд іона; NA — число Авогадро; r — радіус іона; ε0 — електрична стала (ε0 =8,85·10—12 Ф/м). Формула Борна дозволяє визначити величину потенціального бар’єра, який повинен перебороти іон при перетинанні мембрани. Для одновалентних іонів K+ та Na+ ∆W складає від 250 до 350 кДж/моль. Коефіцієнт розподілу, тобто відношення числа іонів, що знаходяться в мембрані, nм до числа іонів, що знаходяться в рідкій фазі, nв розраховується за формулою:
де RT — енергія теплових коливань (R — універсальна газова стала; T
— температура). При температурі 300 К коефіцієнт розподілу складе:
105

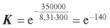 ≈ 10–60.
≈ 10–60.
Тобто з кожних 1060 іонів, що знаходяться у водному розчині, тільки один знаходиться в ліпідній фазі мембрани. Тому імовірність проходження іонів та інших гідрофільних речовин через мембрану внаслідок простої дифузії вкрай мала.
Швидкість перенесення іонів значно зростає, якщо в мембрані існують іонні канали. Відомі, наприклад, канали для іонів калію, натрію, кальцію, хлору. Іонні мембранні канали утворені інтегральними білками. Такий канал може бути або відкритий, або закритий для транспорту іона. Ці два стани реалізуються зміною конформації білків, які формують канали. Зміна конформації, в свою чергу, може бути викликана зміною умов у клітині та позаклітинному середовищі, наприклад, зміною мембранного потенціалу, як у випадку натрієвих і калієвих каналів нервових волокон. Ці канали грають важливу роль при проведенні нервового імпульсу і є найбільше вивченими. Як правило, за допомогою каналів транспортуються гідрофільні речовини, тому пори мають усередині велике число гідрофільних хімічних груп.
Якщо припустити, що електричний потенціал змінюється по всій довжині каналу лінійно (dϕ/dx = ∆ϕ/l = const), відношення концентрацій іонів усередині мембрани біля її меж і в навколишньому середовищі дорівнює коефіцієнту розподілу К, канал однорідний і іони рухаються незалежно один від одного, то транспорт речовин через канали буде описуватися електродифузійним рівнянням Нернста — Планка (5.5).
Однак частіше всього у випадку перенесення речовин через канали більшість цих припущень не виконується, і транспорт описується більш складними рівняннями. Канал, насамперед, неоднорідний за будовою: уздовж його внутрішньої порожнини розташовуються різні хімічні групи, спорідненість іонів до яких неоднакова. Канал може мати один або кілька іонних центрів зв’язування, які, як правило, представляють собою заряджені групи, зв’язавшись із якими, іон потрапляє в потенціальну яму. Для того, щоб потрапити в іншу потенціальну яму іону потрібно перебороти деякий потенціальний бар’єр. Таких бар’єрів уздовж довжини каналу може бути декілька, причому висота їх, як правило, неоднакова. Вона може змінюватися через наявність або відсутність іонів у каналі, зміни трансмембранної різниці потенціалів.
Так як в каналі знаходяться заряджені групи, то припущення про лінійну зміну мембранного потенціалу також не виконується. Це ж може привести і до різних значень коефіцієнта розподілу на різних кінцях каналу.
Багато мембранних каналів настільки вузькі, що іони не можуть у них рухатися в різних напрямках незалежно один від одного: якщо в якійсь
106

потенціальній ямі вже знаходиться іон, то інший може в неї потрапити тільки, якщо буде виштовхнено перший. Це відноситься, наприклад, до калієвих каналів. Зустрічні потоки іонів натрію через натрієві канали можна вважати незалежними, але якщо через ці канали рухаються іони калію, то принцип незалежності вже не виконується.
Для багатьох каналів у залежності від їхньої будови при високих концентраціях іонів у навколишньому середовищі спостерігається ефект насичення (при збільшенні концентрації іонів швидкість їхнього перенесення підвищується, але тільки до певного рівня, вище якого швидкість не змінюється) або блокування (при концентраціях іонів вище деякого значення швидкість перенесення починає знижуватися).
При переході одновалентного іона із навколишнього середовища в канал зміна вільної енергії складає:
де εв — діелектрична проникність води, εк — діелектрична проникність середовища каналу, r — радіус іона, а — радіус каналу. У даному рівнянні перший доданок обумовлений переходом іона з одного середовища в інше, діелектричні проникності яких різняться (див. 5.15), а другий
— зв’язуванням зарядженого іона з протилежно зарядженою групою каналу. У випадку транспорту іона через канал величина ∆W знижується в порівнянні з ∆W переносу безпосередньо через бішар завдяки наявності другого негативного доданку, а також завдяки зменшенню першого доданку внаслідок того, що іон потрапляє в середовище з більшим значенням діелектричної проникності, у порівнянні з діелектричною проникністю мембрани. У результаті дії цих чинників ∆W при переході іона через канал складає біля 30 кДж/моль.
Іонні канали селективні, тобто мають різні величини провідності для різних іонів. Кожний канал транспортує переважно іони одного виду, наприклад, натрієвий канал — іони натрію, калієвий — іони калію. Через катіонні канали не здатні проходити аніони і навпаки. Але для іонів того ж знака, що й основний іон, канал уже не має такої абсолютної селективності. Провідність каналу для інших іонів свого знака нижча, ніж для власного, але відмінна від нуля. Наприклад, специфічні натрієві канали здатні транспортувати також і іони калію, але в 20 разів гірше, ніж іони натрію.
Відповідно до сучасних уявлень, селективність каналу насамперед визначається радіусом іона, що транспортується. Іони, радіус яких перевищує радіус каналу, не здатні проходити через нього внаслідок такої стеричної невідповідності. Іони, що мають менший радіус у порівнянні з радіусом каналу, здавалося б, повинні легко перетинати його. Однак, проникність для них зменшується зі зменшенням радіуса іона. Цей ефект
107

пояснюється тим, що іони у водяному розчині завжди знаходяться в гідратированому стані. Перенесення ж через канал вимагає скидання, якщо не всіх, то принаймні кількох гідратних оболонок, що супроводжується значними витратами енергії. Наприклад, енергія повної гідратації іонів K+ та Na+ складає від 280 кДж/моль до 400 кДж/моль, у той час як енергія теплових коливань при фізіологічних температурах приблизно дорівнює 2,5 кДж/моль. Для транспорту іона через канал потрібні менші витрати енергії, тому що скинута гідратна оболонка іона заміняється іншими полярними групами, у ролі яких виступають фіксовані дипольні групи каналу. Розмір іона повинен бути таким, щоб іон міг взаємодіяти з оптимальною кількістю цих полярних груп. Наприклад, іон калію, потрапивши в селективний фільтр (найбільш вузьке місце в каналі, яке визначає його селективні властивості) калієвого каналу, здатний взаємодіяти зі всіма атомами кисню, розташованими там, у той час як менший за розміром іон натрію, опинившись у калієвому каналі, може зв’язатися тільки з двома атомами кисню. Потенціальна енергія іона калію, який знаходиться в «своєму» каналі, не набагато перевищує його потенціальну енергію у воді, а от зміна потенціальної енергії іона натрію, що потрапив у калієвий канал, вже значна. Таким чином, щонайкраще через канал проникають ті іони, ефективний радіус котрих приблизно дорівнює радіусу каналу.
Селективність каналів може визначатися не тільки залежністю провідності каналу від радіуса іона, що транспортується, але і хімічною будовою іона, як у випадку органічних катіонів. Наприклад, через натрієві канали переносяться іони гідроксиламонію (H3N+–OH) і не переносяться іони метиламонію (H3N+–CH3), хоча їхні радіуси майже однакові і приблизно рівні радіусу каналу. Це пояснюють тим, що гідроксильна група гідроксиламонію здатна утворювати водневі зв’язки із атомами кисню, що вистилають внутрішній діаметр каналу, а метильна група метиламонію з ними не взаємодіє.
Канали можуть блокуватися деякими сполуками, які зв’язуються із селективним фільтром або зі входом у канал, закупорюючи, таким чином, його. Так як молекулаFблокатор повинна стерично відповідати ділянці каналу, що блокується, то для каналів певного виду існують специфічні блокатори. Наприклад, молекули тетрадотоксина блокують натрієві канали, а тетраетіламонію — калієві.
§ 27. Пасивний транспорт речовин за допомогою переносників
Для деяких речовин існують специфічні переносники білкової природи, що знижують енергію активації, необхідну для перенесення речовини через ліпідний бішар. Переносники здійснюють так звану полегшену дифузію. Загальний потік речовин через мембрану при участі пере-
108

носників складається з двох компонент: потоку, викликаного простою дифузією, і потоку, викликаного полегшеною дифузією (рис. 5.2). Тому в даному випадку речовини транспортуються з більшою швидкістю, ніж у випадку простої дифузії. Переносники, що здійснюють пасивний транспорт, переносять речовини із області з більшою концентрацією в область із меншою концентрацією і не вимагають затрат енергії.
Переносники забезпечують транспорт речовин, що самі по собі проникають через мембрану вкрай повільно через високу гідрофільність. Існують переносники для деяких амінокислот, вуглеводів, пуринових і піримідинових основ і нуклеозидів. Таким чином, наприклад, відбувається усмоктування багатьох харчових продуктів у кишечнику, процеси реабсорбції в нирках.
Відмітні ознаки транспорту речовин за допомогою переносників:
1.Переносники мають високу хімічну специфічність, тобто здатні переносити тільки певні субстрати (деякі переносники навіть здатні розрізняти LF і DFізомери тієї самої речовини).
2.Швидкість перенесення зростає пропорційно концентрації речовини, яка переноситься, але лише до деякої певної величини, тобто спостерігається ефект насичення переносника (рис. 5.2, крива 2). Варто мати на увазі, що в реальній клітині навіть при повному насиченні переносника при збільшенні різниці концентрацій ця швидкість все рівно буде зростати за рахунок простої дифузії (рис. 5.2, крива 3).
3.Переносники здатні конкурентно інгібіруватися речовинами, структура яких подібна до структури субстрату.
Потік речовини, яка транспортується переносниками з області, де
їїконцентрація дорівнює с, в область, де концентрація цієї ж речовини дорівнює нулю, описується формулою
(5.16) де Jmax — максимальний потік (потік у випадку насичення переносника); Km — константа спорідненості молекул, що транспортуються, до переносників, чисельно рівна концентрації речовини, при якій J = Jmax/2.
Тоді однобічні потоки речовини з міжклітинного середовища в клітину(Ji) і з клітини назовні (Jo) записуються у вигляді:
де сi і co — концентрації речовини усередині і зовні клітини, відповідно.
109

|
Якщо концентрації речовини по |
|
|
обидва боки мембрани відмінні від нуля, |
|
|
то сумарний потік буде являти собою ал- |
|
|
гебраїчну суму двох різноспрямованих |
|
|
потоків Ji і Jo: |
|
Рис. 5.2. Залежність потоку (J) |
(5.17) |
|
З формули (5.17) видно, що потік |
||
речовини, що переноситься з облас- |
||
ті, де її концентрація дорівнює с, в |
згодом буде зменшуватися, поки кон- |
|
область, де її концентрація дорівнює |
центрації не вирівняються і потік не ста- |
|
нулю, від концентрації с: 1 — проста |
не рівним нулю. Вище уже вказувалося, |
|
дифузія; 2 — транспорт за участі |
що таке вирівнювання в організмі, як |
|
переносника; 3 — сумарна швидкість |
||
переносу. |
правило, не настає внаслідок того, що |
|
|
речовини увесь час утворюються або |
|
|
витрачаються в біохімічних реакціях. |
Якщо концентрація якоїсь речовини з одного боку мембрани мала у порівнянні з її концентрацією на протилежному боці, то величину потоку з деяким наближенням можна визначати по рівнянню (5.16).
Графік рівняння (5.16), поданий на рис. 5.2, незручний для визначення констант Jmax і Km. За допомогою нескладних перетворень (5.16) можна звести до вигляду (рівняння Лайнуївера — Берка):
Тоді графік залежності 1/J = f(1/с) становитиме пряму (рис. 5.3). Отримавши експериментальні значення 1/J і 1/с на деякому проміжку прямої, можна шляхом екстраполяції визначити значення Jmax і Km на перетині прямої з осями координат. Цей метод дозволяє також визначити присутність конкурентного інгібітора переносу даної речовини в системі (рис. 5.3, пряма 2).
Одним з найбільш вивчених випадків пасивного транспорту речовин за допомогою переносників є перенесення глюкози в еритроцити. Переносник DFглюкози здатний транспортувати також речовини, що структурно нагадують субстрат, наприклад, DFманозу, DFксилозу, DFарабінозу і DFгалактозу, але не відповідні LFцукри. Перелічені DFізомери вуглеводів викликають конкурентне інгібірування переносника DFглюкози, що змушений переносити ці речовини замість свого власного субстрату. Інший конкурентний інгібітор транспорту глюкози в деякі клітини — флоридзин1 — також конкурентно зв’язується з переносником, але в клітину не проникає.
Як видно, для того, щоб специфічний переносник цукру переніс
110