

1.3.2. Преобразование энергии раздражителя в рецепторах
В результате действия адекватного раздражителя у большинства рецепторов увеличивается проницаемость клеточной мембраны для катионов, что приводит к току катионов в клетку и деполяризации ее мембраны. Лишь в фотопигментных клетках сетчатки в связи с особенностями управления ионными каналами (имеется темновой ток натрия в клетку) вслед за поглощением фотонов света мембранный потенциал увеличивается с — 20—40 мВ до приблизительно — 70 мВ, то есть происходит гиперполяризация (подробнее см. в главе 3).
Изменение величины мембранного потенциала рецепторов в ответ на действие стимула представляет собой рецеп-торный потенциал — входной сигнал первичных сенсорных нейронов. Если величина рецепторного потенциала достигнет критического уровня деполяризации или превысит его, генерируются потенциалы действия, с помощью которых сенсорные нейроны передают информацию о действующих стимулах в центральную нервную систему (рис. 1.2).
Генерация потенциалов действия происходит в ближайшем к рецепторам перехвате Ранвье миелинизированных волокон или ближайшей к рецепторам части мембраны без-миелинового волокна. Минимальная сила адекватного стимула, достаточная для генерации потенциалов действия в
16

Рис. 1.2. Зависимость между силой раздражителя, величиной реценторного потенциала и частотой потенциалов действия
При действии подпорогового стимула 1 возникает подпоро-говый рецепторный потенциал, потенциалы действия не образуются. При действии порогового стимула 2 и при пороговом значении потенциала образуются потенциалы действия. При действии надпорогового стимула 3 частота потенциалов действия возрастает. При проведении потенциалов действия по нервному волокну частота потенциалов действия не изменяется.
первичном сенсорном нейроне, определяется как его абсолютный порог. Минимальный прирост силы стимула, сопровождающийся значимым изменением реакции сенсорного нейрона, представляет собой дифференциальный порог его чувствительности.
Информация о силе действующего на рецепторы стимула кодируется двумя способами: частотой потенциалов действия, возникающих в сенсорном нейроне (частотное кодирование), и числом сенсорных нейронов, возбудившихся в ответ на действие стимула. При увеличении силы действующего на рецепторы раздражителя повышается амплитуда рецеп-торного потенциала, что, как правило, сопровождается увеличением частоты потенциалов действия в сенсорном нейроне первого порядка. Чем шире имеющийся частотный диапазон потенциалов действия у сенсорных нейронов, тем большее число промежуточных значений силы раздражителя способна различать сенсорная система. Первичные сенсор-
17
ные нейроны одинаковой модальности различаются порогом возбуждения, поэтому при действии слабых стимулов возбуждаются только наиболее чувствительные нейроны, но с увеличением силы раздражителя на него реагируют и менее чувствительные нейроны, имеющие более высокий порог раздражения. Чем больше первичных сенсорных нейронов возбудится одновременно, тем сильнее будет их совместное действие на общий нейрон второго порядка, что в итоге отразится на субъективной оценке интенсивности действующего раздражителя.
Длительность ощущения зависит от реального времени между началом и прекращением воздействия на рецепторы, а также от их способности уменьшать или даже прекращать генерацию нервных импульсов при продолжительном действии адекватного стимула. При длительном действии стимула порог чувствительности рецепторов к нему может повышаться, что определяется как адаптация рецепторов. Механизмы адаптации не одинаковы в рецепторах разных модальностей, среди них различают быстро адаптирующиеся (например, тактильные рецепторы кожи) и медленно адаптирующиеся рецепторы (например, проприоцепторы мышц и сухожилий). Быстро адаптирующиеся рецепторы сильнее возбуждаются в ответ на быстрое нарастание интенсивности стимула (физический ответ), а их быстрая адаптация способствует освобождению восприятия от биологически незначительной информации (например, контакт между кожей и одеждой). Возбуждение медленно адаптирующихся рецепторов мало зависит от скорости изменения стимула и сохраняется при его длительном действии (тонический ответ), поэтому, например, медленная адаптация про-приоцепторов позволяет человеку получать нужную ему для сохранения позы информацию в течение всего необходимого времени.
Существуют сенсорные нейроны, генерирующие потенциалы действия спонтанно, то есть при отсутствии раздражения (например, сенсорные нейроны вестибулярной системы), такая активность называется фоновой. Частота нервных импульсов в этих нейронах может увеличиваться или уменьшаться в зависимости от интенсивности действующего на вторичные рецепторы стимула, кроме того, она может определяться на-
18
правлением, в котором отклоняются чувствительные волоски механорецепторов. Например, отклонение волосков вторичных механорецепторов в одну сторону сопровождается повышением фоновой активности сенсорного нейрона, которому они принадлежат, а в противоположную сторону — понижением его фоновой активности. Указанный способ рецепции позволяет получать информацию и об интенсивности стимула, и о направлении, в котором он действует.
1.4. Рецептивные поля
Рецептивным полем называется область, занимаемая совокупностью всех рецепторов, стимуляция которых приводит к возбуждению сенсорного нейрона (рис. 1.3). Макси-
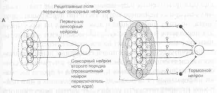
Рис. 1.3. Рецептивные поля первичных сенсорных нейронов и сенсорных нейронов второго порядка
А. Рецептивные поля первичных сенсорных нейронов ограничены областью распределения их чувствительных окончаний. Рецептивное поле переключательного нейрона образуется из суммы рецептивных полей конвергирующих к нему первичных сенсорных нейронов.
Б. Раздражение центральной или периферической областей рецептивного поля сенсорного нейрона второго и следующих порядков дает противоположный эффект. Как видно на схеме, раздражение центра рецептивного поля вызовет возбуждение проекционного нейрона, а раздражение периферической области — торможение с помощью интернейронов переключательного ядра (латеральное торможение).
В результате контраста, создаваемого между центром и периферией рецептивного поля, облегчается восприятие раздражителя.
19
Величина рецептивных полей сенсорных нейронов второго и следующих порядков больше, чем у первичных сенсорных нейронов, поскольку центральные нейроны получают информацию от нескольких конвергирующих к ним нейронов предшествующего уровня. От центра рецептивного поля информация передается непосредственно к сенсорным нейронам следующего порядка, а от периферии — к тормозным интернейронам переключательного ядра, поэтому центр и периферия рецептивного поля являются реципрок-ными по отношению друг к другу. В результате сигналы от центра рецептивного поля беспрепятственно достигают следующего иерархического уровня сенсорной системы, тогда как сигналы, поступающие от периферии рецептивного поля, тормозятся (в другом варианте организации рецептивного поля легче пропускаются сигналы от периферии, а не от центра). Такая функциональная организация рецептивных полей обеспечивает выделение наиболее значимых сигналов, легко различаемых на контрастном с ними фоне.
20
1.5. Переработка информации в переключательных ядрах
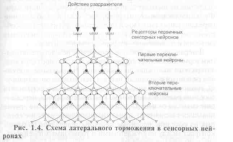
Сенсорные нейроны низшего иерархического уровня способны передавать электрические сигналы одновременно нескольким нейронам следующего уровня благодаря дивергенции к ним своих аксонов. Это повышает надежность передачи информации от одного иерархического уровня к другому и позволяет сохранять чувствительность сенсорной системы при утрате отдельных нейронов. Возбуждение нейронов более высокого иерархического уровня определяется не только действием на них нейронов предыдущего уровня, но и механизмом латерального торможения, существующего в переключательных ядрах (рис. 1.4).
Тормозные нейроны (их тела выделены черным цветом) возбуждаются возвратными коллатералями переключательных сенсорных нейронов: чем сильнее возбужден переключательный нейрон, тем большее торможение соседних переключательных нейронов он вызовет. Степень возбуждения переключательных нейронов определяется интенсивностью раздражения центра или периферии его рецептивного поля. В результате возбуждение будет передаваться от одной части рецептивного поля на фоне торможения или прерывания передачи сигнала от другой ею части, что создает эффект контрастирования, способствующий различению действующих стимулов.
21
Каждое переключательное ядро представляет собой, во-первых, распределительную систему, образующую проекции на другие структуры мозга для передачи им информации. Во-вторых, в любом переключательном ядре происходит переработка информации, заключающаяся в изменениях параметров электрических сигналов. Переключательные ядра образованы разными типами нервных клеток, которые, в зависимости от выполняемой функций, подразделяются на два класса: проекционные {релейные) и локальные интернейроны.
Проекционные нейроны имеют достаточно длинные аксоны, с помощью которых переносят электрические сигналы от одного переключательного ядра к другому, передавая информацию на следующий уровень ее переработки. Короткие аксоны возбуждающих и тормозных локальных интернейронов не выходят за пределы переключательного ядра. Функция локальных интернейронов состоит в облегчении передачи одних сигналов и подавлении других, что способствует передаче на следующий уровень наиболее значимых сигналов и подавлению «информационного шума».
Латеральное торможение происходит вследствие активации тормозных интернейронов переключательного ядра кол -латералями возбуждающих нейронов. Чем сильнее возбужден релейный сенсорный нейрон, тем больше он активирует тормозные нейроны, которые подавляют активность соседних релейных нейронов. Релейные нейроны, передающие сигналы от центра рецептивного поля, сильнее других повышают активность тормозных нейронов, влияние которых на соседние релейные нейроны делает еще большим контраст между возбужденными и тормозимыми нейронами, выделяя непрерывающуюся линию передачи избранных сигналов.
1.5.1. Нисходящее торможение и усиление в сенсорных системах
Высшие уровни сенсорной системы регулируют переработку информации в низших по отношению к ним переключательных ядрах посредством нисходящего торможения или усиления передаваемых сигналов. Нисходящее торможение (рис. 1.5) происходит вследствие активации тормозных
22
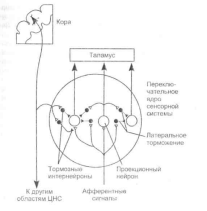
Рис. 1.5. Схема нисходящего торможения в сенсорных системах
Передачу афферентных сигналов от одного иерархического уровня сенсорной системы к другому в переключательных ядрах регулируют высшие иерархические уровни. На схеме показан принцип нисходящего торможения коры, которое осуществляется с помощью тормозных интернейронов (выделены черным цветом) переключательного ядра. Возбуждение тормозных ин-тернейронов под влиянием коры препятствует передаче афферентных сигналов в переключательном ядре.
нейронов переключательного ядра, деятельность которых определяют нисходящие пути, начинающиеся на более высоком иерархическом уровне сенсорной системы. В результате нисходящего торможения повышается порог афферентной синаптической передачи в переключательном ядре низшего уровня. Нисходящее торможение представляет собой регуляцию сенсорного восприятия по механизму отрица-
23
тельной обратной связи. Тормозные интернейроны переключательных ядер уменьшают частоту передаваемых на следующий уровень сигналов, в связи с чем сенсорное ощущение ослабляется.
Нисходящее усиление происходит с помощью возбуждающих интернейронов переключательного ядра, которые активируются аксонами нейронов более высокого иерархического уровня. Вследствие этого в управляемом по механизму положительной обратной связи переключательном ядре понижается порог синаптической передачи, что делает возможным прохождение относительно слабых сигналов на следующий иерархический уровень. Одновременное использование механизмов отрицательной и положительной обратной связи позволяет освобождаться от избыточной информации, подавлять «шум», то есть нейронную активность, не содержащую физиологически значимых сообщений, и одновременно выделять и усиливать сигналы, на которых будет сосредоточено внимание.