

Свойства высокомолекулярных соединений
Макромолекулярная структура полимеров обусловливает появление целого ряда особенностей свойств этих веществ. Целый ряд свойств проявляется у полимеров иначе, нежели у низкомолекулярных соединений; кроме того, появляются новые свойства, которыми в принципе не могут обладать низкомолекулярные соединения.
В этом разделе будут рассмотрены: А. Особенности физикохимии полимеров – как в конденсированной фазе, так и в растворах. Б. Особенности химических свойств полимеров.
Особенности физикохимии полимеров
В основе практически всех особенностей физических свойств полимеров (да в большой степени и химических) лежит пространственная структура макромолекул – расположение макромолекул в пространстве и возможность его изменения в ходе теплового движения или под действием внешнего поля. Поэтому вначале имеет смысл остановиться на рассмотрении пространственной организации макромолекулярных тел.
Пространственная структура макромолекул.
Свойства высокомолекулярных соединений в огромной степени зависят от пространственной структуры их макромолекул и от характера изменений этой структуры. Использование ВМС в качестве пластмасс, эластомеров (каучуков и резин) или волокон напрямую связано с различными типами пространственной организации их макромолекул и с разными возможностями ее изменения под действием внешних сил. В частности, в пластмассах пространственная структура макромолекул «заморожена», и при действии механических нагрузок изменяются лишь длины связей и валентные углы, т.е. пластмассы проявляют свойства твердых тел. В эластических полимерах (эластомерах) пространственная структура легко и весьма значительно изменяется при действии механических нагрузок и столь же легко восстанавливается при снятии нагрузок; это приводит к появлению больших обратимых деформаций. В волокнах макромолекулы вытянуты и взаимно ориентированы, что приводит к анизотропии механических свойств, в частности, к повышенной прочности вдоль оси ориентации. Сложнейшие и высокоспецифические биологические функции белков и нуклеиновых кислот возможны только при условии специфической, причем строго определенной, пространственной организации макромолекул этих биополимеров.
В свою очередь, пространственная структура макромолекул зависит: а). От химического строения; б). От внешних факторов, прежде всего от температуры, от молекулярного окружения полимера (например, в случае растворов - от растворителя), от приложенного внешнего поля (механической нагрузки, электромагнитного поля).
Как известно, пространственное расположение любых ковалентно построенных молекул определяется, во-первых, величинами валентных углов, во-вторых, пространственной конфигурацией (если возможна стереоизомерия), в-третьих, конформациями (если возможно вращение вокруг простых связей). Большинство наиболее известных полимеров имеют сравнимые величины валентных углов, и на специфике пространственной структуры полимеров в зависимости от величин этих углов можно специально не останавливаться. Конфигурации звеньев играют весьма важную роль при формировании пространственной структуры макромолекул полимеров, в звеньях которых возможна стереоизомерия. В частности цис-полиизопрен (натуральный каучук) и транс-полиизопрен (гуттаперча) кардинально различаются по своим механическим свойствам. Как уже упоминалось выше, стереорегулярные полимеры могут образовывать более регулярно построенные пространственные структуры, что исключительно важно для биополимеров – все они являются стереорегулярными (изотактическими).
Но особо важное значение в формировании пространственной структуры макромолекул всех полимеров играют конформации полимерных цепей; на их роли остановимся более подробно.
Как известно, конформации молекулы – это варианты расположения ее атомов в пространстве, которые могут переходить друг в друга путем вращения вокруг одинарных связей. Естественно, чем больше таких связей в молекуле, тем большее число конформаций теоретически возможно; число возможных конформаций любой достаточно большой полимерной цепи невообразимо велико; соответственно исключительно велико разнообразие пространственных форм, в которых может находиться полимерная цепь.
Для характеристики конформаций используют величины двугранных углов между связями, соседними с той связью, вокруг которой происходит вращение (эти углы также называют торсионными). Например, на рис.10 представлены наиболее характерные конформации (I-VI) для бутана, 1СН3-2СН2-3СН2-4СН3, возникающие при вращении вокруг связи 2С–3С; углы характеризуют взаимное расположение связей 1С-СН3 и 2С-СН3:
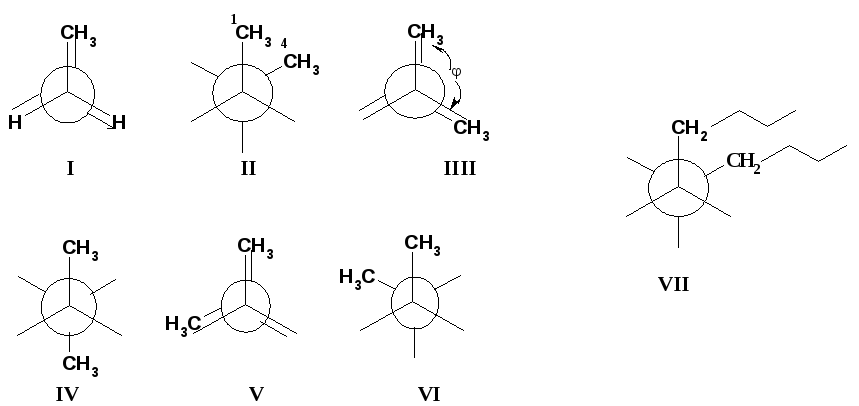
Рис.10. Конформации н-бутана (I-VI) и полиэтилена (VII).
Для конформации I угол равен 00, для II = +600 , для III , для IV, для V= -120o, для VI= -600 [согласно рекомендациям IUPAC торсионный угол имеет знак плюс, если для того, чтобы перейти к заслоненному положению однотипных групп (на рис.2 это группы СН3) ближнюю группу нужно повернуть по часовой стрелке, и знак минус - если против часовой стрелки]
Аналогичный подход используется для характеристики конформаций полимерных цепей. Так, в простейшем случае полиэтилена конформации полимерных цепей характеризуются двугранными углами между фрагментами цепи –СН2~ , присоединенными к той или иной связи (рис. 2, VII).
Для характеристики конформаций биополимеров используют специфические обозначения. В частности для обозначения конформаций полипептидных цепей (рис.11) используют величины торсионных углов исвязанных с вращением соответственновокруг связей N-CH(R) и связей СН(R)-СО(вокруг пептидной связи вращение при обычных температурах не происходит вследствие того, что она имеет характер частично двойной связи). Например, угол описывает взаимное расположение связей N4-С3 и С5-С6, а угол i - связей С5-N4 и С6-N7.
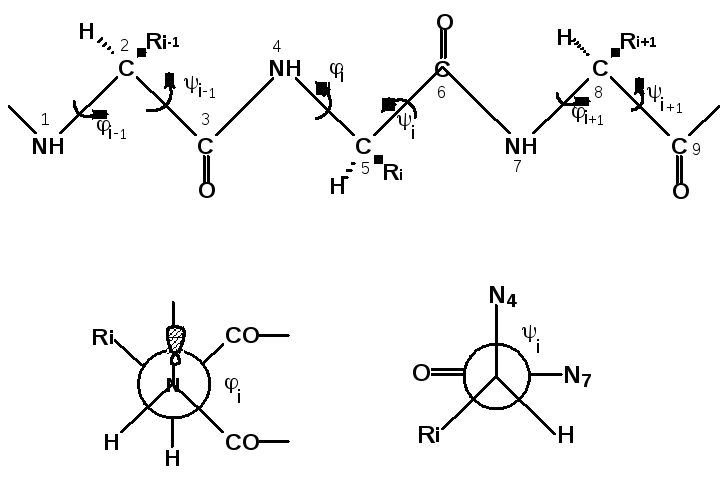
Рис.11. Торсионные углы, характеризующие конформации
полипептидных цепей.
Конформации полинуклеотидных цепей описываются величинами торсионных углов (рис.12).
O P O C H 2 O O - O O 1 ' 2 ' 3 ' 4 ' 5 ' N N N N N H 2 N N H N N O N H 2 A B 1 2 3 4 5 6 7 8 9 
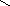
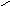
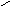
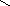
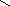







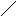





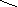

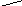









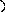









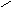












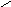















Рис.12. Торсионные углы, характеризующие конформации
полинуклеотидных цепей.
Для полисахаридов возможности вращения вокруг простых связей существенно меньше: если гликозидная связь образована с участием вторичного гидроксила, например, по типу 1-4’ (рис. 13, А), то вращение возможно вокруг двух связей (углы и); если она образована по схеме 1-6’ (рис. 13, В) – вокруг трех связей.
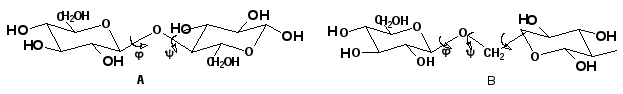
Рис.13. Возможности внутреннего вращения в полисахаридах.
Любая трехмерная структура полимерной цепи может быть описана набором величин торсионных углов, относящихся ко всем связям этой цепи, вокруг которых возможно внутреннее вращение.
Для определения точной пространственной структуры всей полимерной молекулы необходимо учитывать не только конформацию цепи, но и конформации, связанные с поворотами боковых заместителей вокруг связей, соединяющих их с основной цепью. Например, при описании полной пространственной структуры нуклеиновых кислот учитывают также конформации, связанные с поворотом гетероциклических оснований вокруг N-гликозидной связи; им соответствуют торсионные углы (рис.4); в частности, для пуриновых оснований такими углами принято считать двугранные углы между связями С1-О и N9-С8; в двойной спирали ДНК пуриновые основания находятся в анти-конформации относительно дезоксирибозного цикла (это хорошо видно на рис.4 справа), =0.
Аналогично для описания полной конформации полипептидных цепей нужно учесть конформации, связанные с поворотом боковых групп Ri.
Хотя в целом ряде случаев учет пространственного расположения боковых групп достаточно важен, все же наибольшее значение имеют конформации полимерных цепей: они наиболее тесно связаны с практически важными свойствами синтетических и искусственных полимеров и биологическими функциями биополимеров.