

364 :: 365 :: 366 :: 367 :: 368 :: 369 :: Содержание
297 :: Содержание

ОБМЕН УГЛЕВОДОВ
Углеводы входят в состав живых организмов и вместе с белками, липидами и нуклеиновыми кислотами определяют специфичность их строения и функционирования. К углеводам относят соединения, обладающие разнообразными и зачастую сильно отличающимися функциями. Углеводы участвуют во многих метаболических процессах, но прежде всего они являются основными поставщиками энергии. На долю углеводов приходится примерно 75% массы пищевого суточного рациона и более 50% от суточного количества необходимых калорий. Однако неправильно сводить функцию углеводов только к энергетическому обеспечению процессов жизнедеятельности организма. Следует отметить и структурную роль углеводов. Так, в виде гликозаминогликанов углеводы входят в состав межклеточного матрикса. Большое число белков (ферменты, белки-транспортёры, белки-рецепторы, гормоны) - гликопротеины, углеводная составляющая которых повышает их специфичность. Например, различия в строении олигосахаридных фрагментов клеточной оболочки эритроцитов обеспечивают групповую принадлежность крови. Из углеводов в процессе метаболизма образуется большое число органических соединений, которые служат исходными субстратами для синтеза липидов, аминокислот, нуклеотидов. Производные углеводов - глюкурониды - участвуют в детоксикации ксенобиотиков и инактивации веществ эндогенного происхождения. Углеводы могут быть синтезированы в организме с использованием других метаболитов: некоторых аминокислот, глицерина, молочной кислоты. Углеводы нельзя считать незаменимыми компонентами пищи. Однако если исключить углеводы из диеты, то следствием может быть гипогликемия, для компенсации которой будут расходоваться белки и липиды. Таким образом, углеводы - обязательные пищевые компоненты, потому что помимо их основной энергетической функции (клеточные "дрова") углеводы участвуют во многих метаболических клеточных процессах.
297
297 :: Содержание
298 :: 299 :: 300 :: 301 :: 302 :: 303 :: 304 :: 305 :: Содержание
I. Строение углеводов
Термин "углеводы", предложенный в XIX столетии, был основан на предположении, что все углеводы содержат 2 компонента - углерод и воду, и их элементарный состав можно выразить общей формулой Cm(H2O)n. Хотя из этого правила есть исключения и оно не абсолютно точно, тем не менее указанное определение позволяет наиболее просто характеризовать класс углеводов в целом. К тому же попытка, предпринятая Комиссией по химической номенклатуре, заменить термин "углеводы" на "глициды" не удалась. Новый термин не получил широкого признания. Термин "углеводы" укоренился и общепризнан.
Углеводы можно разделить на 3 основные группы в зависимости от количества составляющих их мономеров: моносахариды, олигосахариды и полисахариды.
А. Моносахариды
Моносахариды - производные многоатомных спиртов, содержащие карбонильную группу. В зависимости от положения в молекуле карбонильной группы моносахариды подразделяют на альдозы и кетозы.
Альдозы содержат функциональную альдегидную группу -НС=О, тогда как кетозы содержат кетонную группу >С=О. Название моносахарида зависит от числа составляющих его углеродных атомов, например альдотриозы, кетотриозы, альдогексозы, кетогексозы и т.д.
Моносахариды по строению можно отнести к простым углеводам, так как они не гидролизуются при переваривании, в отличие от сложных, которые при гидролизе распадаются с образованием простых углеводов. Строение основных представителей моносахаридов показано на рис. 7-1.
В пище человека (фрукты, мёд, соки) содержится небольшое количество моносахаридов, в основном глюкоза и фруктоза.
Глюкозаявляется альдогексозой. Она может существовать в линейной и циклической формах. Циклическая форма глюкозы, предпочтительная в термодинамическом отношении, обусловливает химические свойства глюкозы. Как и все гексозы, глюкоза имеет 4 асимметричных углеродных атома, обусловливающих наличие сте-реоизомеров. Возможно образование 16 стереоизомеров, наиболее важные из которых D- и L-глюкоза. Эти типы изомеров зеркально отображают друг друга (рис. 7-2).
Расположение Н- и ОН-групп относительно пятого углеродного атома определяет принадлежность глюкозы к D- или L-ряду. В организме млекопитающих моносахариды находятся в D-конфигурации, так как к этой форме глюкозы специфичны ферменты, катализирующие её превращения. В растворе при образовании циклической формы моносахарида образуются ещё 2 изомера (α- и β-изомеры), называемые аномерами, обозначающие определённую конформа-цию Н- и ОН-групп относительно С, (рис. 7-3). У α-D-глюкозы ОН-группа располагается ниже плоскости кольца, а у β-D-глюкозы, наоборот, над плоскостью кольца.
Фруктоза является кетогексозой (кетогругша находится у второго углеродного атома). Фруктоза так же, как и глюкоза, существует в циклической форме, образуя α- и β-аномеры (рис. 7-4).
Б. Реакции моносахаридов
Присутствие гидроксильных, альдегидных и кетонных групп позволяет моносахаридам вступать в реакции, характерные для спиртов, альдегидов или кетонов. Эти реакции довольно многочисленны. В данном разделе будут описаны лишь некоторые из них, причём в основном имеющие наибольшее биологическое значение.
В этом разделе основные реакции моносахаридов рассмотрены на примере D-глюкозы (рис. 7-5), хотя надо иметь в виду, что в метаболизме углеводов принимают участие и другие моносахариды, а также их производные.
Мутаротация, или аномеризация - взаимопревращение аномерных форм моносахаридов, α- и β-формы аномеров находятся в растворе в состоянии равновесия. При достижении этого равновесия происходит мутаротация - размыкание и замыкание пиранового кольца и, соответственно, изменение расположения Н- и ОН-групп при первом углероде моносахарида.
Образование гликозидов. Гликозидная связь имеет важное биологическое значение, потому что именно с помощью этой связи осуществляется ковалентное связывание моносахаридов в составе олиго- и полисахаридов. При образовании гликозидной связи аномерная ОН-группа одного моносахарида взаимодействует с ОН-группой другого
298
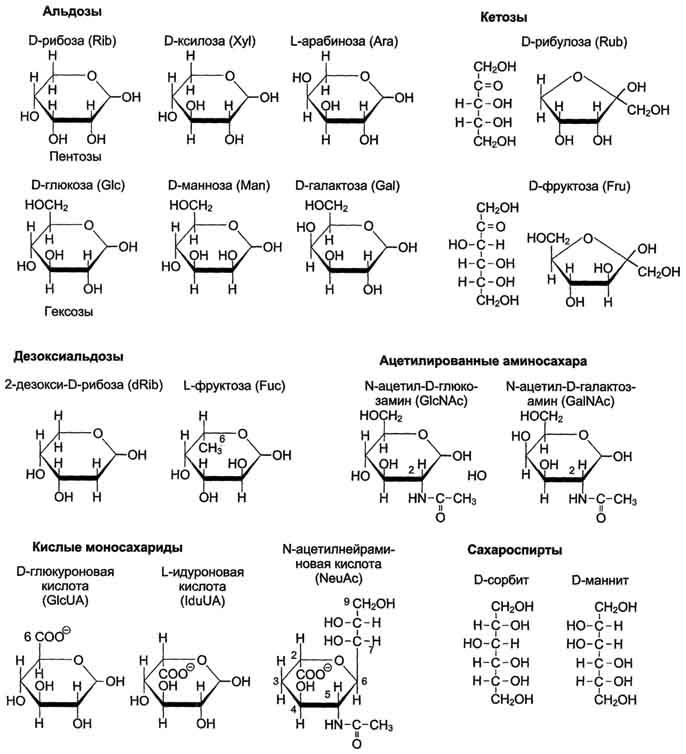
Рис. 7-1. Важнейшие моносахариды.
299
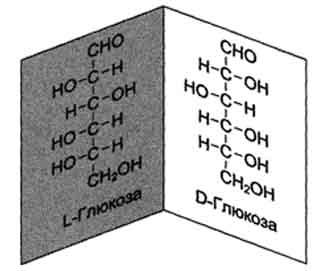
Рис. 7-2. D- и L-изомеры глюкозы.
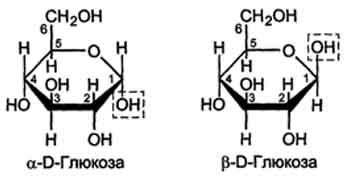
Рис. 7-3. α- и β-аномеры D-глюкозы.
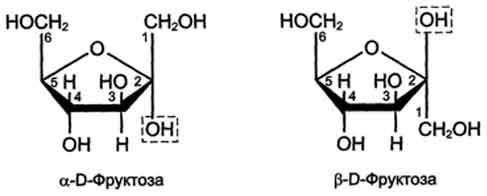
Рис. 7-4. α- иβ-аномеры D-фруктозы.
300
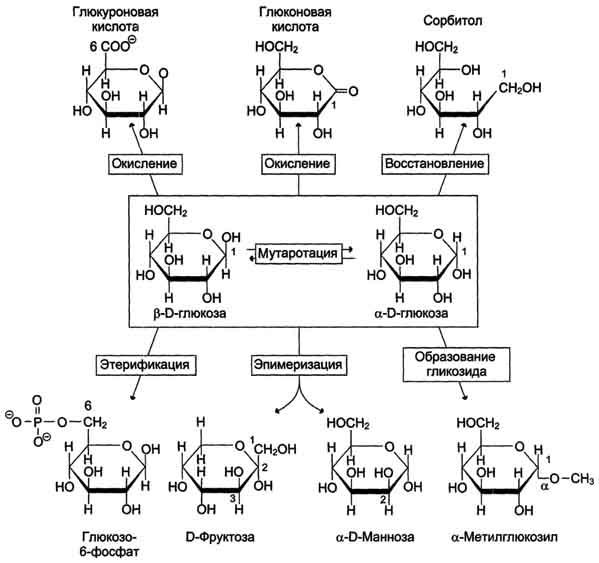
Рис. 7-5. Реакции моносахаридов.
моносахарида или спирта. При этом происходят отщепление молекулы воды и образование О-гликозидной связи. Все линейные олигомеры (кроме дисахаридов) или полимеры содержат мономерные остатки, участвующие в образовании двух гликозидных связей, кроме концевых остатков, образующих только одну гликозидную связь. Некоторые гликозидные остатки могут образовывать три гликозидные связи, что характерно для разветвлённых олиго- и полисахаридов. Олиго- и полисахариды могут иметь концевой остаток моносахарида со свободной аномерной ОН-группой, не использованной при образовании гликозидной связи. В этом случае при размыкании цикла возможно образование свободной карбонильной группы, способной окисляться. Такие олиго- и полисахариды обладают восстанавливающими свойствами и поэтому называются восстанавливающими, или редуцирующими (рис. 7-6).
Аномерная ОН-группа моносахарида может взаимодействовать с NН2-группой других соединений, что приводит к образованию N-гликозидной связи. Подобная связь присутствует в нуклеотидах и гликопротеинах (рис. 7-7).
Этерификация. Это реакция образования эфирной связи между ОН-группами моносахаридов и различными кислотами. В метаболизме углеводов важную роль играют фосфоэфиры - эфиры моносахаридов и фосфорной кислоты. В метаболизме глюкозы особое место
301
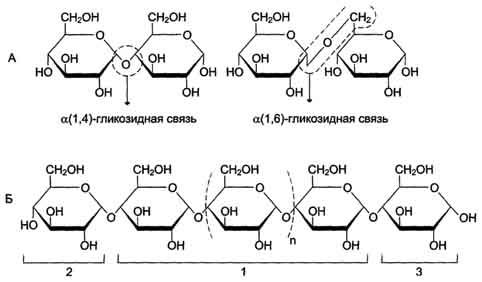
Рис. 7-6. Строение полисахарида. A.Образование α-1,4- и α-1,6-гликозидных связей.Б.Строение линейного полисахарида: 1 - α-1,4-гликозидные связи между мономерами; 2 - невосстанавливающий конец (невозможно образование свободной карбонильной группы у аномерного углерода); 3 - восстанавливающий конец (возможно размыкание цикла с образованием свободной карбонильной группы у аномерного углерода).
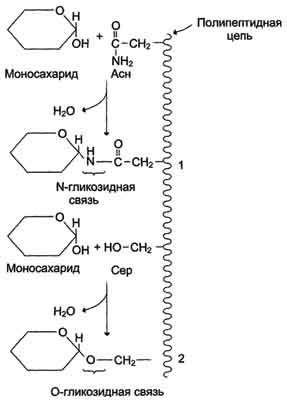
Рис. 7-7. Образование О- и N-гликозидных связей в гликопротеинах.1 - N-гликозидная связь между амидной группой аспарагина и ОН-группой моносахарида; 2 - О-гликозидная связь между ОН-группой серина и ОН-группой моносахарида.
302
занимает глюкозо-6-фосфат. Образование глюкозо-6-фосфата происходит в ходе АТФ-зависимой реакции при участии ферментов, относящихся к группе киназ. АТФ в данной реакции выступает как донор фосфатной группы. Фосфоэфиры моносахаридов могут образовываться и без использования АТФ. Например, глюкозо-1-фосфат образуется из гликогена при участии Н3РО4. Физиологическое значение фос-фоэфиров моносахаридов заключается в том, что они представляют собой метаболически активные структуры. Реакция фосфорилирования моносахаридов важна для метаболизма ещё и потому, что клеточная мембрана мало проницаема для этих соединений, т.е. клетка удерживает моносахариды благодаря тому, что они находятся в фосфорилированной форме.
Окисление и восстановление. При окислении концевых групп глюкозы -СНО и -СН2ОН образуются 3 различных производных. При окислении группы -СНО образуется глюконовая кислота. Если окислению подвергается концевая группа -СН2ОН, образуется глюкуроновая кислота. А если окисляются обе концевые группы, то образуется сахарная кислота, содержащая 2 карбоксильные группы. Восстановление первого углерода приводит к образованию сахароспир-та - сорбитола.
В. Олигосахариды
Олигосахариды содержат несколько (от двух до десяти) остатков моносахаридов, соединённых гликозидной связью. Дисахариды - наиболее распространённые олигомерные углеводы, встречающиеся в свободной форме, т.е. не связанной с другими соединениями. По химической природе дисахариды представляют собой гликозиды, которые содержат 2 моносахарида, соединённые гликозидной связью в α- или β-конфигурации. В пище содержатся в основном такие дисахариды, как сахароза, лактоза и мальтоза (рис. 7-8).
Сахароза - дисахарид, состоящий из α-D-глюкозы и β-D-фруктозы, соединённых α,β-1,2-гликозидной связью. В сахарозе обе аномерные ОН-группы остатков глюкозы и фруктозы участвуют в образовании гликозидной связи. Следовательно, сахароза не относится к восстанавливающим сахарам. Сахароза - растворимый дисахарид со сладким вкусом. Источником сахарозы служат растения, особенно сахарная свёкла, сахарный тростник. Последнее объясняет возникновение тривиального названия сахарозы - "тростниковый сахар".
Лактоза - молочный сахар; важнейший дисахарид молока млекопитающих. В коровьем молоке содержится до 5% лактозы, в женском молоке - до 8%. В лактозе аномерная ОН-группа первого углеродного атома остатка D-галактозы связана β-гликозидной связью с четвёртым углеродным атомом D-глюкозы (β-1,4-связь). Поскольку аномерный атом углерода остатка глюкозы не участвует в образовании гликозидной связи, следовательно, лактоза относится к восстанавливающим сахарам.
Мальтоза поступает с продуктами, содержащими частично гидролизованный крахмал, например, солод, пиво. Мальтоза также образуется при расщеплении крахмала в кишечнике. Мальтоза состоит из двух остатков D-глюкозы, соединённых α-1,4-гликозидной связью.
Изомальтоза - промежуточный продукт, образующийся при расщеплении крахмала в кишечнике. Состоит из двух остатков D-глюкозы, но соединены эти моносахариды α-1,6-гликозидной связью.
Г. Полисахариды
Структурные различия между полисахаридами определяются:
строением моносахаридов, составляющих цепь;
типом гликозидных связей, соединяющих мономеры в цепи;
последовательностью остатков моносахаридов в цепи.
В зависимости от строения остатков моносахаридов полисахариды можно разделить на гомополисахариды (все мономеры идентичны) игетерополисахариды (мономеры различны). Оба типа полисахаридов могут иметь как линейное расположение мономеров, так и разветвлённое.
В зависимости от выполняемых ими функций полисахариды можно разделить на 3 основные группы:
резервные полисахариды, выполняющие энергетическую функцию. Эти полисахариды служат источником глюкозы, используемым организмом по мере необходимости. Резервная функция этих углеводов обеспечивается их полимерной природой. Полисахариды
303
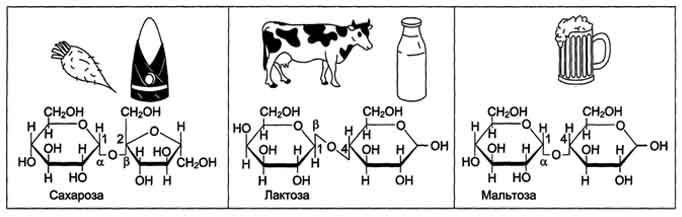
Рис. 7-8. Дисахариды пищи.
менее растворимы, чем моносахариды, следовательно они не влияют на осмотическое давление и поэтому могут накапливаться в клетке, например, крахмал - в клетках растений, гликоген - в клетках животных;
структурные полисахариды, обеспечивающие клеткам и органам механическую прочность (см. раздел 15);
полисахариды, входящие в состав межклеточного матрикса, принимают участие в образовании тканей, а также в пролиферации и дифференцировке клеток. Полисахариды межклеточного матрикса водорастворимы и сильно гидратированы (см. раздел 15).
В пище человека в основном содержатся полисахариды растительного происхождения - крахмал, целлюлоза. В меньшем количестве поступает полисахарид животных - гликоген.
Крахмал - наиболее важный углеводный компонент пищевого рациона. Это резервный полисахарид растений, содержащийся в наибольшем количестве (до 45% от массы сухого вещества) в зёрнах злаков (пшеница, кукуруза, рис и др.), а также луковицах, стеблях и клубнях растений (в картофеле примерно 65%). Крахмал - разветвлённый полисахарид, состоящий из остатков глюкозы (гомогликан). Он находится в клетках растений в виде гранул, практически нерастворим в воде.
Крахмал состоит из амилозы и амилопектина (рис. 7-9). Амилоза - неразветвлённый полисахарид, включающий 200-300 остатков глюкозы, связанных α-1,4-гликозидной связью. Благодаря α-конфигурации глюкозного остатка, полисахаридная цепь имеет конформацию спирали. Синяя окраска при добавлении йода к раствору крахмала обусловлена наличием такой спирали. Амилопектин имеет разветвлённую структуру. В местах ветвления остатки глюкозы соединены α-1,6-гликозидными связями. Линейные участки содержат примерно 20-25 остатков глюкозы. При этом формируется древовидная структура, в которой имеется лишь одна аномерная ОН-группа. Крахмал - высокомолекулярное соединение, включающее сотни тысяч остатков глюкозы. Его молекулярная масса составляет порядка 105-108Д.
Целлюлоза (клетчатка) - основной структурный полисахарид растений. Это самое распространённое органическое соединение на земле. Доля целлюлозы в клеточных стенках растений составляет 40-50%. Целлюлоза имеет молекулярную массу порядка 106Д, длина молекулы может доходить до 6-8 мкм.
Целлюлоза - линейный полисахарид гомогликан, построенный из остатков глюкозы, соединённых между собой β-1,4-гликозидными связями. Пищеварительная система человека не имеет ферментов, гидролизующих β-связи в полисахаридах. Поэтому целлюлоза - неиспользуемый углевод, но этот пищевой компонент необходим для нормального протекания переваривания.
Гликоген - полисахарид животных и человека. Так же, как крахмал в растениях, гликоген в клетках животных выполняет резервную функцию, но, так как в пище содержится лишь небольшое количество гликогена, он не имеет пищевого значения.
304
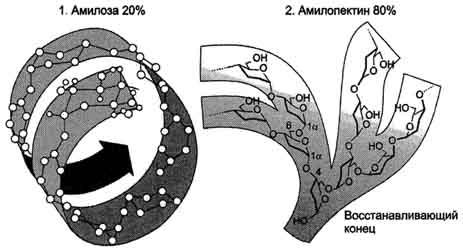
Рис. 7-9. Строение крахмала.
Гликоген представляет собой структурный аналог крахмала, но имеет большую степень ветвления: примерно на каждые 10 остатков глюкозы приходится одна α-1,6-гликозидная связь.
305