

МИНИСТЕРСТВО ЗДРАВООХРАНЕНИЯ И СОЦИАЛЬНОЙ ЗАЩИТЫ НАСЕЛЕНИЯ
РЕСПУБЛИКИ ТАДЖИКИСТАН
ТАДЖИКСКИЙ ГОСУДАРСТВЕННЫЙ МЕДИЦИНСКИЙ УНИВЕРСИТЕТ ИМ. АБУАЛИ ИБНИ СИНО
Шукуров Ф.А., Халимова Ф.Т.
КРАТКИЙ КУРС
НОРМАЛЬНОЙ ФИЗИОЛОГИИ
ДУШАНБЕ - 2017
УДК 612
Ш-95
Рецензенты:
Ишанкулова Б.А. - доктор медицинских наук , профессор кафедры фармакологии Таджикского государственного медицинского университета имени Абуали ибни Сино
Устоев М.Б. - д.б.н., профессор, заведующий кафедрой физиологии человека
и животных Таджикского национального университета
Вахобов А.А. – кандидат биологических наук, доцент, заведующий кафедрой
анатомии и физиолгии Таджикского государственного педагогического
университета имени С.Айни
Учебник рекомендован к печати Министерством народного образования и науки РТ от 23.11.2017 за № 17/26
Шукуров Ф.А.
Ш-95 Краткий курс нормальной физиологии: учебник /Ф.А. Шукуров, Ф.Т. Халимова. – МБОО «Олами китоб», 2017. – 160с
Учебник предназначен для улучшения подготовки студентов к практическим занятиям и другим формам контроля знаний по изучаемой дисциплине. Нормальная физиология как наука о жизнедеятельности здорового человека и физиологических основ здорового образа жизни, является методологическим фундаментом медицины, главным образом его профилактического направления. Нормальная физиология является также научной основой диагностики здоровья и прогнозирования функциональной активности организма человека. Стратегическое направление врача общей практики должно быть направлено на предупреждение болезней человека, на охрану и поддержание здоровья и здорового образа жизни, а не только на лечение болезней.
Нормальная физиология является завершающей учебной дисциплиной в разделе доклинического медико-биологического образования студентов, которая органически связана с биологией, биофизикой, биохимией, анатомией, гистологией с одной стороны и с патологической физиологией, фармакологией и пропедевтическими клиническими дисциплинами с другой стороны. В представленном кратком курсе систематизированы основные термины, определения и понятия, используемые в курсе по нормальной физиологии с учетом последних достижений в этой области.
Материал учебника излагается кратко, доходчиво и методически грамотно. Данное издание адресовано студентам высших медицинских учебных заведений, обучающимся на медицинском, педиатрическом,стоматологическом и фармацевтическом факультетах, на факультете общественного здравоохраненияапо, а также на биологических факультетах.
Физиология – это научный стержень,
на котором держатся все медицинские науки.
К. Бернар
Общая физиология
Понятия о раздражителях, раздражимости, раздражении, возбудимости и биологических реакциях. На ткани организма постоянно действуют какие-то факторы, которые могут находиться внутри или вне организма, которые называются раздражителями. Раздражители – это факторы внешней или внутренней среды, обладающие запасом энергии и при действии которых на ткань отмечается их биологическая реакция. Различают два вида биологических реакций: специфические и неспецифические. Специфические реакции характерны для какой-то строго определенной ткани. Например, специфическая реакция мышечной ткани – это сокращение, для железистой ткани – это выделение секрета или инкрета (гормона), для нервной ткани – это генерация и передача нервного импульса. Таким образом, специфической деятельностью обладают специализированные ткани. Неспецифические реакции характерны для любой живой ткани. Например, изменение интенсивности обмена веществ, изменение мембранного потенциала покоя, изменение ионного градиента и т.д. Теперь дадим определение таким понятиям, как раздражение, раздражимость, возбудимость и биологические реакции. Раздражение – это любые воздействия на ткань. В ответ на раздражения возникают биологические реакции ткани, т.е. ответная реакция ткани на действие раздражителя. Все биологические реакции делятся на две группы: 1) неспецифические реакции – эти реакции свойственны для любой ткани и проявляются они в виде изменения интенсивности обмена веществ, величины мембранного потенциала покоя и др.; 2) специфические реакции – эти реакции свойственны только для специализированных тканей: для мышечной ткани - это сокращение, для железистой ткани – это выделение секрета или инкрета (гормона), для нервной ткани – это генерация (образование) нервного импульса и передача его к рабочему органу (эффектору). Раздражимость – это универсальное свойство живой материи и отражает способность любой живой ткани изменять свою неспецифическую деятельность под влиянием раздражения. Возбудимость – это свойство специализированных тканей и отражает способность ткани реагировать на раздражение изменением своих специфических реакций. Возбудимость ткани определяется его пороговой силой: чем меньше пороговая сила, тем больше возбудимость ткани.
Классификация раздражителей (по силе и биологическому соответствию). Все многообразие раздражителей можно выделить в отдельные группы. Классификация раздражителей зависит от того, что берется за основу:
По биологическому соответствию, то есть насколько раздражитель соответствует данной ткани. По данной классификации различают два вида: 1) адекватные – раздражители, которые соответствуют данной ткани. Например, для сетчатки глаза свет – все остальные раздражители не соответствуют сетчатке, для мышечной ткани – нервный импульс и т.д.; 2) неадекватные – раздражители, которые не соответствуют данной ткани. Для сетчатки глаза все раздражители кроме светового будут неадекватные, а для мышечной ткани все раздражители, кроме нервного импульса. Для сетчатки глаза все раздражители кроме светового будут неадекватные, а для мышечной ткани все раздражители, кроме нервного импульса. Адекватный раздражитель для каждой ткани только один, а неадекватных раздражителей – много. Пороговая сила адекватного раздражителя меньше, чем пороговая сила неадекватного раздражителя.
По силе – различают пять основных раздражителей (рис.1): 1) подпороговые раздражители – это сила раздражителя при которой не возникает ответная реакция (локальный ответ, т.е. изменения происходят только на месте действия раздражителя); 2) пороговый раздражитель – это минимальная сила, которая вызывает ответную реакцию при бесконечном времени действия. Эту силу еще называют реобазой – она единственная для каждой ткани; 3)надпороговые, или субмаксимальные; 4) максимальный раздражитель – это минимальная сила при которой возникает максимальная ответная реакция ткани; 5) сверхмаксимальные раздражители – при этих раздражителях реакция ткани либо максимальная, либо уменьшается, либо временно исчезает. Таким образом, для каждой ткани существует один пороговый раздражитель, один максимальный и множество подпороговых, надпороговых и сверхмаксимальных.

Рис. 1 Зависимость ответной реакции ткани от силы раздражителя: А - величина ответной реакции, Б – сила раздражителя
Функциональные состояния ткани, их физиологическая характеристика. Различают три функциональных состояний ткани: покой, возбуждение и торможение. Состояние покоя – это пассивный процесс, при котором отсутствуют внешне выраженные проявления специфической деятельности. Например, сокращение, секреция и др. Состояние возбуждения и торможения – это активные процессы, при которых в одном случае усиливается специфическая деятельность ткани (возбуждение), а в другом – либо полностью исчезает проявление специфической деятельности, либо уменьшается, хотя на ткань при этом продолжает действовать раздражитель (торможение).
Возбудимость, мера возбудимости (пороговая сила и хронаксия). Возбудимость – это свойство специализированных тканей и отражает способность ткани реагировать на раздражение изменением своих специфических реакций. Возбудимость ткани определяется его пороговой силой и хронаксией. Пороговая сила – это минимальная сила, которая вызывает ответную реакцию при бесконечном времени действия. Эту силу еще называют реобазой – она единственная для каждой ткани: пороговая сила обратнопропорциональна возбудимости: чем больше пороговая сила, тем меньше возбудимость ткани. Хронаксия – это минимальное время, в течении которого необходимо действовать силой в две реобазы, чтобы получить ответную реакцию ткани. Для определения хронаксии необходимо вначале определить пороговую силу (реобазу), затем удвоить эту силы (взять две реобазы) и найти минимальное время, которое необходимо, чтобы при действии силы в две реобазы произошла ответная реакция ткани. Для определения хронаксии используется прибор - хронаксиметр. Этот прибор широко используется в клинике нервных болезней для определения хронаксии нервов и мышц, чтобы оценить возбудимость этих тканей и тем самым определить степень поражения этих тканей с одной стороны и с другой стороны – определить эффективность назначенного лечения. Если после проведенного лечения хронаксия не уменьшается, значит лечение было не правильным.
Законы раздражения возбудимых тканей: силы, времени. Законы раздражения возбудимых тканей: 1) закон силы – возбудимость обратно-пропорциональна пороговой силе: чем больше пороговая сила, тем меньше возбудимость. Однако для возникновения возбуждения недостаточно только действия силы раздражения. Необходимо, чтобы это раздражение длилось какое-то время; 2) закон времени действия раздражителя. При действии одной и той же силы на разные ткани потребуется разная длительность раздражения, что зависит от способности данной ткани к проявлению своей специфической деятельности, то есть возбудимости: наименьшее время потребуется для ткани с высокой возбудимостью и наибольшее время - с низкой возбудимостью. Таким образом, возбудимость обратно-пропорциональна времени действия раздражителя: чем меньше время действия раздражителя, тем больше возбудимость.
При взаимоотношении живого объекта со средой практически не проявляются только закон силы или только закон времени. Любое воздействие на биологические системы одновременно обладает двумя неотделимыми друг от друга свойствами – силой и длительностью действия. При этом усиление раздражения приводит к укорочению времени, необходимого для возбуждения, но не беспредельно, то есть для каждой силы раздражения существует свой порог времени (минимальное время). Таким образом, сила и длительность раздражения находятся в обратной зависимости, что отражается кривой «силы-времени».
Закон скорости нарастания силы раздражителя. Аккомодация. В сложных взаимоотношениях организма со средой ему приходится сталкиваться не только с изменением силы и длительности действия раздражителя, но и с быстротой их изменений. Быстроту изменения силы характеризует скорость нарастания силы раздражения, что отражается третьим законом – зависимости возбудимости ткани от скорости нарастания силы раздражителя: чем меньше пороговая скорость нарастания силы раздражителя, тем больше возбудимость ткани. Для каждой ткани существует своя пороговая скорость нарастания силы раздражения. Если мы опустим лапку лягушки в горячую воду ( +50о), то возникает ответная реакция в виде отдергивания лапки. Теперь опустим лапку лягушки в воду с комнатной температурой - никакой реакции не будет. Не вынимая лапки лягушки, медленно будем нагревать воду до +50о – реакции не будет. Таким образом, реакция ткани зависит не только от силы раздражителя, но и от скорости нарастания силы раздражения. Если на ткань действовать силой, скорость нарастания которой ниже пороговой величины, то ткань не реагирует на этот раздражитель. Такое явление получило название аккомодация, то есть приспособление ткани на действие раздражителя. Сущность аккомодации заключается в повышении порога возбудимости, которое возникает при медленно нарастающей силе раздражения. При этом уменьшается критический уровень деполяризации, что приводит к увеличению пороговой силы, уменьшению возбудимости и в связи с этим ткань не реагирует на данный раздражитель.
Кривая силы-времени. Показать на этой кривой реобазу, хронаксию, полезное время. Эта кривая (рис.2) отражает зависимость силы раздражителя (F) от времени его действия (t). Эта зависимость в пределах А-В обратно-пропорциональна: чем больше силараздражителя, тем меньше время его действия. На данной кривой можно определить следующие показатели: пороговую силу, или реобазу (ОС), полезное время (ОР) и хронаксию (ON). Последовательность действий (алгоритм), необходимых для определения хронаксии на кривой силы-времени: 1) находим точку на кривой силы-времени, после которой кривая параллельна оси времени – t (В), 2) от этой точки проводим прямую параллельно оси времени до пересечения с осью силы – F (С), 3) находим пороговую силу (ОС), 4) удвоим пороговую силу (ОМ), 5) от точки М проводим прямую параллельную оси времени до пересечения с кривой (Е), 6) из точки Е опускаем перпендикуляр на ось времени (N), 7) находим хронаксию (ON) – минимальное время, которое необходимо, чтобы при действии силы в две реобазы произошла ответная реакция ткани. Хронаксия так же, как и реобаза, является мерой возбудимости: чем больше хронаксия, тем меньше возбудимость ткани.
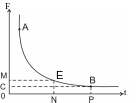
Рис. 2 Кривая силы-времени
Биопотенциалы, их виды. МПП и МПД, величина и способ регистрации. Биопотенциалы – это разность потенциалов между двумя точками живой ткани, отражающая ее биоэлектрическую активность. Можно сказать, что биопотенциалы – это электрический ток в живых системах.
Различают три вида биопотенциала:
1) мембранный потенциал покоя (МПП);
2) мембранный потенциал действия (МПД);
3) потенциал градиента основного обмена (метаболический потенциал).
МПП – это разность потенциалов между наружней и внутренней поверхностью мембраны в состоянии покоя. При этом на наружней поверхности мембраны скапливаются положительные заряды, а на внутренней поверхности – отрицательные заряды. Величина МПП соответствует –70 - -90 мв. Знак минус означает, что в покое на внутренней поверхности мембраны скапливаются отрицательные заряды. Такое распределение зарядов для клеточной мембраны характерно, когда клетка находится в состоянии физиологического покоя. Мембрана с расположенными отрицательными зарядами на внутренней ее поверхности и положительными на наружней находится в состоянии поляризации. Регистрация МПП осуществляется только внутриклеточным способом. Для этой цели используют микроэлектроды – их диаметр должен соответствовать порам мембраны. Использование микроэлектрода связана с тем, что мембрана при этом практически не повреждается при введении электрода во внутрь клетки.
МПД – это разность потенциалов между наружней и внутренней поверхностью мембраны при возбуждении. Можно дать другое определение: МПД – это разность между возбужденной и невозбужденной частью мембраны. Таким образом, МПД можно регистрировать двумя способами: 1) внутриклеточным – для этого также используют микроэлектроды; 2) внеклеточным способом. Этот способ возможен благодаря тому, что возбуждение не сразу охватывает всю мембрану клетки, а распространяется волнообразно. При этом на наружной поверхности возбужденной части мембраны скапливаются отрицательные заряды, а на невозбужденной части – положительные. Благодаря внеклеточной регистрации, МПД широко используется в клинике: а) ЭМГ – электромиография, регистрация МПД скелетных мышц, используется для изучения функционального состояния мышц; б) ЭЭГ – электроэнцефалография, регистрация МПД мозга, используется для изучения функционального состояния коры больших полушарий; в) ЭГГ – электрогастрография, регистрация МПД желудка, используется для изучения двигательной функции желудка; г) ЭКГ – электрокардиограмма, регистрация МПД сердечной мышцы, используется для изучения функционального состояния сердечной мышцы. Величина МПД соответствует +110 +120 мв. Знак плюс означает, что при возбуждении внутренняя поверхность мембраны заряжена положительно. Мембрана с расположенными положительными зарядами на внутренней ее поверхности и отрицательными на наружней находится в состоянии деполяризации.
Механизм возникновения мембранного потенциала покоя. В настоящее время экспериментально доказано, что большую роль в таком распределении зарядов играют ионы натрия и калия, которые неравномерно распределены во внутриклеточной и внеклеточной среде. В клетках возбудимых тканей концентрация ионов калия достигает 150 ммоль/л, а во внеклеточной среде – 4-5 ммоль/л, то есть концентрация ионов калия во внутриклеточной среде в 30-50 раз больше, чем во внеклеточной среде. Концентрация ионов натрия, наоборот, больше во внеклеточной среде, чем во внутриклеточной (во внеклеточной среде ионов натрия 140 ммоль/л, а во внутриклеточной среде – 14 ммоль/л, то есть во внеклеточной среде ионов натрия в 10 раз больше. В механизме возникновения МПП большую роль играют специальные поры – каналы, просветы которых так узки, что проходить через них способны только относительно мелкие ионы натрия и калия. Причем, в мембране имеются специальные каналы, пропускающие калий (калиевые каналы), или натрий (натриевые каналы). Известно, что в состоянии покоя поры мембраны, пропускающие ионы калия (калиевые каналы), в основном открыты, а поры, пропускающие ионы натрия (натриевые каналы), в основном закрыты. В связи с этим в состоянии покоя проницаемость для ионов калия (пассивный транспорт по градиенту) в несколько раз больше, чем для ионов натрия. Каждый выходящий ион калия остается на наружней поверхности мембраны, так как анион не может пройти через калиевый канал и остается на внутренней поверхности мембраны. В состоянии покоя происходит поляризация мембраны: на наружней поверхности мембраны скапливаются положительные заряды (ионы калия), а на внутренней поверхности – анионы (отрицательные заряды). Следует отметить, что в состоянии покоя натриевые каналы в основном закрыты, поэтому они в очень незначительном количестве проникают во внутриклеточную среду. Причем, каждый зашедший натрий захватывает анион с внутренней поверхности мембраны и проходит во внутриклеточную среду. В связи с этим ион калия, находящийся на наружной поверхности мембраны, также, соединяясь с анионом, проходит во внеклеточную среду. Таким образом, каждый зашедший во внутрь клетки ион натрия, уменьшает поляризацию, а, следовательно, уменьшает величину МПП, а каждый, вышедший из клетки ион калия, увеличивает поляризацию и величину МПП. Наличие МПП в клетке сваидетельствует о готовности мембраны реагировать на раздражение.
Механизм возникновения мбранного потенциала действия. В настоящее время наибольшее признание получила ионная теория, объясняющая механизм возникновения МПД. Экспериментально доказано, что большую роль в таком распределении зарядов играют ионы натрия и калия, которые неравномерно распределены во внутриклеточной и внеклеточной среде. В клетках возбудимых тканей концентрация ионов калия достигает 150 ммоль/л, а во внеклеточной среде – 4-5 ммоль/л, то есть концентрация ионов калия во внутриклеточной среде в 30-50 раз больше, чем во внеклеточной среде. Концентрация ионов натрия, наоборот, больше во внеклеточной среде, чем во внутриклеточной (во внеклеточной среде ионов натрия 140 ммоль/л, а во внутриклеточной среде – 14 ммоль/л, то есть во внеклеточной среде ионов натрия в 10 раз больше. В механизме возникновения МПД большую роль играют специальные поры – каналы, просветы которых так узки, что проходить через них способны только относительно мелкие ионы натрия и калия. Известно, что при действии раздражителя изменяются свойства мембраны: поры (каналы), пропускающие ионы натрия открываются, а поры, пропускающие ионы калия закрываются. В связи с этим после нанесения раздражения резко увеличивается проницаемость мембраны для ионов натрия и натрий начинает по градиенту (пассивный транспорт) заходить в клетку. Каждый заходящий ион натрия остается на внутренней поверхности мембраны, так как анион не может пройти через натриевый канал и остается на наружней поверхности мембраны. В состоянии возбуждения происходит деполяризация мембраны: на наружней поверхности мембраны скапливаются отрицательные заряды (анионы), а на внутренней поверхности – положительные (ионы натрия). Таким образом, каждый зашедший во внутрь клетки ион натрия, уменьшает поляризацию, а, следовательно, уменьшает величину МПП, что приводит к деполяризации мембраны – возникает МПД
Фазы МПД, механизм возникновения каждого периода. При раздражении клетки изменяются свойства мембраны таким образом, что открываются натриевые каналы, а калиевые закрываются. В связи с этим ионы натрия начинают заходить в клетку, так как их вне клетки в 10 раз больше. Начинается перезарядка мембраны – деполяризация. В момент раздражения наружная поверхность мембраны имеет положительные заряды, поэтому первые порции ионов натрия заходят в клетку медленно, так как при этом действует электростатическая отталкивающая сила. (рис.3)

Рис.3 Фазы мембранного потенциала действия
Медленный заход натрия (б) происходит до определенного уменьшения МПП (до –50-70 мВ), после чего перестает действовать отталкивающая сила. Этот уровень МПП, при котором отсутствует отталкивающая сила, называется критический уровень деполяризации (КУД). Уменьшение МПП до КУД соответствует первой фазе МПД – порог деполяризации (А). От величины КУД зависит пороговая сила, а, следовательно, и возбудимость ткани (чем меньше КУД, тем больше пороговая сила и меньше возбудимость ткани). При уменьшении МПП до уровня КУД начинается залповый заход натрия в клетку (в) и МПП резко уменьшается за счет перезарядки мембраны – это вторая фаза МПД – пик МПД (В). Эта фаза имеет два периода: а) период деполяризации (в) – в этот период происходит резкое уменьшение МПП до определенного уровня (+20+30 мВ). Во время этого периода мембрана теряет свой нормальный заряд (свою «поляризацию»), поэтому этот период называют периодом деполяризации. Как правило, деполяризация переходит за нулевую линию и внутренняя поверхность мембраны приобретает положительный заряд – эта положительная часть пика МПД называется овершут. б) период реполяризации (д) – после достижения своего максимального уровня происходит восстановление исходной величины МПП, то есть мембрана возвращается в состояние поляризации. При этом до уровня КУД происходит быстрое восстановление, затем реполяризация замедляется. В реполяризационной части пика МПД большую роль играет повышение проводимости для инов калия. Кроме этого, реполяризации мембраны способствует работа натриевого насоса, благодаря чему происходит выкачивание ионов натрия из клетки, чтобы вернуть прежнее соотношение ионов натрия внутри и вне клетки (восстановить прежнюю асимметрию ионов натрия). Период реполяризации осуществляется до уровня КУД, затем начинается третья фаза МПД – следовой потенциал (С). Эта фаза также состоит из двух периодов: а) период следовой деполяризации (е) – при этом реполяризация еще не достигает уровня МПП; б) период следовой гиперполяризации (ж) – при этом величина МПП становится больше, чем было до раздражения. Это происходит, потому что реполяризация осуществляется за счет работы натриевого насоса и за счет увеличения проницаемости ионов калия – в результате этого ионов калия выходит больше и количество положительных зарядов на наружной поверхности мембраны становится больше (гиперполяризация). Для предотвращения ионного равновесия в определенный момент срабатывает механизм калиевого насоса, и калий начинает заходить в клетку (транспорт ионов калия против градиента) состояние мембраны возвращается к исходной поляризации. Длительность МПД составляет около 1мс в нервах, 10 мс в скелетной мышце и более 200 мс в миокарде.
Соотношение фаз МПД с фазами возбудимости. Если нормальную возбудимость (в условиях физиологического покоя) принять за 100%, то в ходе одного цикла возбуждения она изменяется как в сторону повышения (экзальтации), так и в сторону понижения (рефрактерности). Изменение возбудимости ткани в разные фазы МПД отражено на рисунке 4. В первую фазу МПД – порога деполяризации (А) - возбудимость ткани возрастает – становится выше нормы (более 100%) эта фаза возбудимости называется супернормальная возбудимость, или фаза экзальтации. Такая же повышенная возбудимость в период следовой деполяризации (С е). В период деполяризации пика МПД (В в) возбудимость полностью исчезает (0%) – эта фаза абсолютной рефрактерности. В период реполяризации пика МПД возбудимость снижена (меньше 100%) – эта фаза относительной рефрактерности (В д). Такое же снижение возбудимости отмечается в период следовой гиперполяризации (С ж). Таким образом, при возбуждении возбудимость ткани неоднородна: может повышаться (фаза экзальтации) в фазу порога деполяризации и в период следовой деполяризации МПД, может уменьшаться (фаза относительной рефрактерности) в период реполяризации пика МПД и в период следовой гиперполяризации, может полностью отсутствовать (фаза абсолютной рефрактерности) в период деполяризации пика МПД.
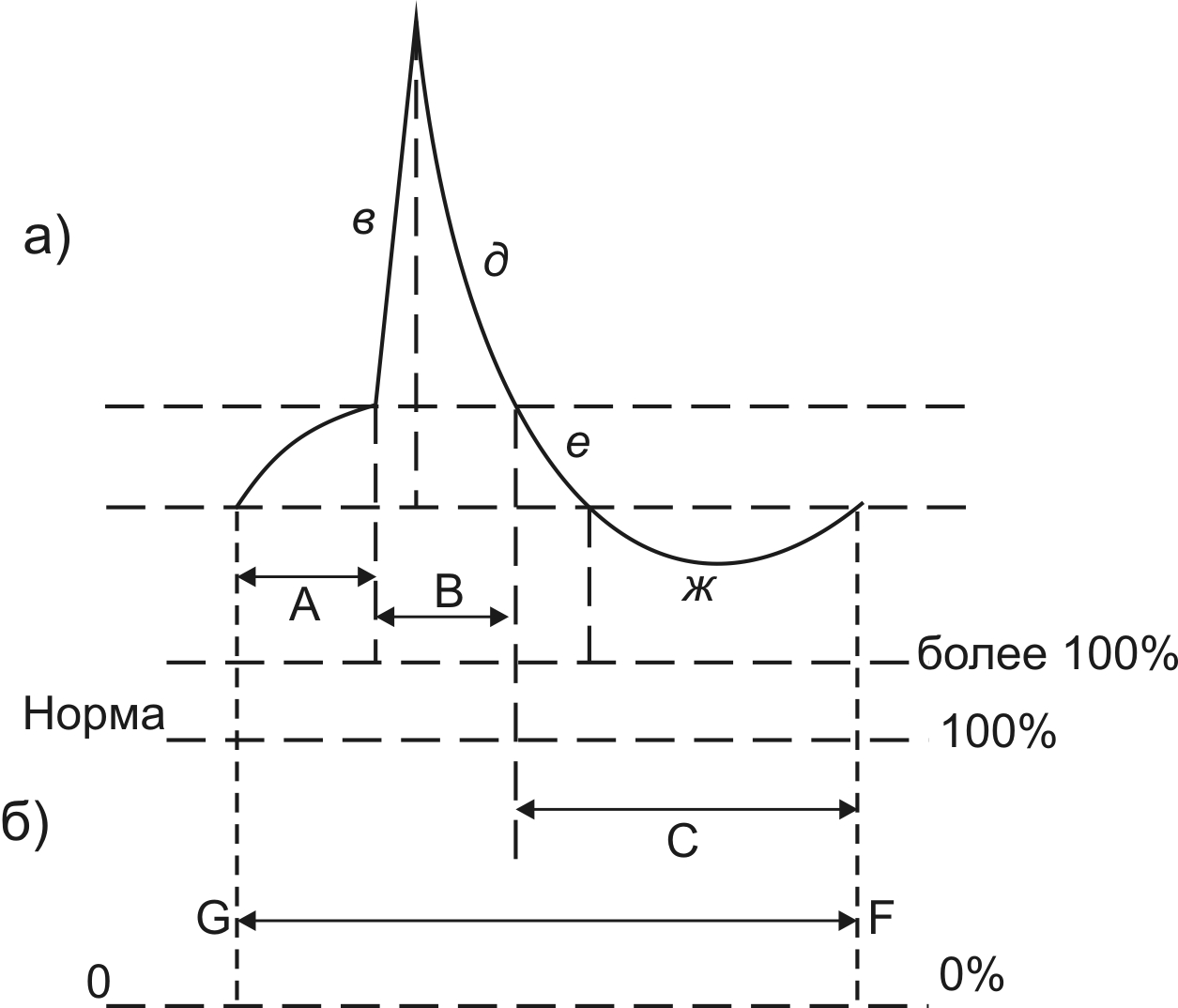
Рис. 4. Изменение возбудимости в разные фазы МПД
Биологическое значение абсолютной рефрактерности заключается в том, что эта фаза защищает живую ткань от чрезмерных энергетических трат и позволяет «без помех» осуществлять текущую приспособительную реакцию.
Активный и пассивный транспорт ионов через мембрану. Калий-натриевый насос, его роль в возникновении МПП и МПД, механизм работы. Выход ионов калия из клетки (при возниковении МПП) и заход ионов натрия внутрь клетки (при возникновении МПД) осуществляется пассивно – по концентрационному градиенту, то есть из области высокой концентрации ионы проникают в область низкой концентрации. Однако, наличие только пассивного транспорта привело бы к исчезновению разности концентраций, что несовместимо в нормальной ткани, то есть исчезновение разности концентраций ионов приводит к гибели клетки (жизнь – это ионная ассиметрия клетки). Следовательно, должен существовать еще один ток ионов через мембрану, поддерживающий ионную асимметрию клетки на каком-то оптимальном уровне. Этот ток ионов осуществляется активно, против градиента (калий-натриевый насос): при этом ионы калия будут заходить в клетку за счет калиевого насоса, который начинает функционировать при сильном увеличении МПП (следовая гиперполяризация), а ионы натрия – выходить из клетки – за счет натриевого насоса, который начинает функционировать при максимальном значении МПД (овершут). Таким образом, калий-натриевый насос это условный механизм, который обеспечивает активный транспорт ионов калия (калиевый насос) и натрия (натриевый насос). В каждой клетке имеется свое пороговое соотношение ионов калия и натрия. Когда в клетке возникает это пороговое соотношение ионов калия и натрия, происходит активация аденозинтрифосфатазы мембраны и происходит расщепление АТФ, освобождается энергия, которая обеспечивает активный транспорт ионов калия и натрия – эта энергия и есть калий-натриевый насос. За счет этой энергии восстанавливается прежнее соотношение ионов калия и натрия внутри и вне клетки. Этот насос будет функционировать до оптимального соотношения ионов калия и натрия во внеклеточной и внутриклеточной среде.
Мышцы, их классификация и свойства. Типы скелетных мышц, их характеристика. Двигательная единица. Мышечные волокна делятся на две группы: 1)поперечно-полосатые мышцы, которые, в свою очередь делятся на два вида: а) скелетные мышцы, б) сердечная мышца (миокард); 2) гладкие мышцы. По международной номенклатуре скелетные мышцы делятся на три типа: 1) I тип мышечных волокон (быстрые мышцы, белые, окислительные, ааэробные). Эти мышцы практически не содержат миоглобина, поэтому сокращаются в безкислородной среде. Энергия для сокращения этих мышц образуется за счет окисления глюкозы, поэтому их еще называют аэробные, или окислительные; IIА тип мышечных волокон (медленные, красные, окислительно-гликолитичес-кие, аэробно-анаэробные); IIВ тип мышечных волокон (красные, медленные, анаэробные, гликолитические). IIА и IIВ типы мышечных волокон содержат много миоглобина, поэтому имеют красный цвет. IIВ тип мышечных волокон отличаются от IIА тем, что первые хорошо васкулизированны (содержат много капилляр, имеют хорошее кровоснабжение). По своей функции скелетные мышцы можно разделить на: 1) фазные – эти мышцы генерирут МПД, который распространяется по всей мышце, они быстро сокращаются и быстро расслабляются (отмечается четкая фазность мышечного сокращения); 2) тонические – эти мышцы не способны генерировать полноценный МПД, распространяющегося типа, поэтому эти мышцы сокращаются и длительно находятся в этом состоянии (в этих мышцах отсутствует четкая фазность в процессе сокращения)
По своей локализации скелетные мышцы различают: 1)интрафузальные – они локализованны в рецепторах мышц (мышечном веретене); 2) экстрафузальные – все мышечные волокна, принадлежащие данной мышце и не входящие в состав мышечного веретена.
![]()
Рис. 5. Двигательная единица
Двигательная единица (рис. 5) состоит из трех элементов: Мотонейрона (1), локализованного в передних рогах спинного мозга; эфферентного нерва (2) и скелетной мышцы (3).
Режимы сокращения мышц. Выделяют три основных режима сокращения мышц (рис. 6):

А Б В Г
Рис. 6. Режимы мышечного сокращения
Изотонический режим (В) – при этом сокращение мышцы происходит за счет укорочения мышечного волокна, а напряжение (тонус) при этом практически не меняется (в2 меньше, чем в), поперечное сечение не изменяется (а=а2); изометрический режим – (Б) - при этом сокращение мышц происходит за счет увеличения напряжения (тонуса), а длина мышечных волокон практически не меняется - увеличивается поперечное сечение мышцы а1 больше, чем а: длина мышцы не изменяется (в=в1); ауксотонический режим, или смешанный режим сокращения (Г). При этом происходит укрочение мышцы (в3 меньше в) и увеличение поперечного сечения (тонуса): а3 больше а.
Структура мышечного волокна и протофибриллы. Мышечное волокно состоит из множества миофибрилл, каждая из которых состоит из большого количества протофибрилл и каждая протофибрилла состоит из миофиламентов. Таким образом, структурно-функциональная единица мышцы – миофиламенты. На поперечном разрезе (рис. 7) видно, что каждая миофиламента состоит из одного толстого волокна (миозина - 1) и вокруг него шесть тонких нитей (волокон) – актина (2)
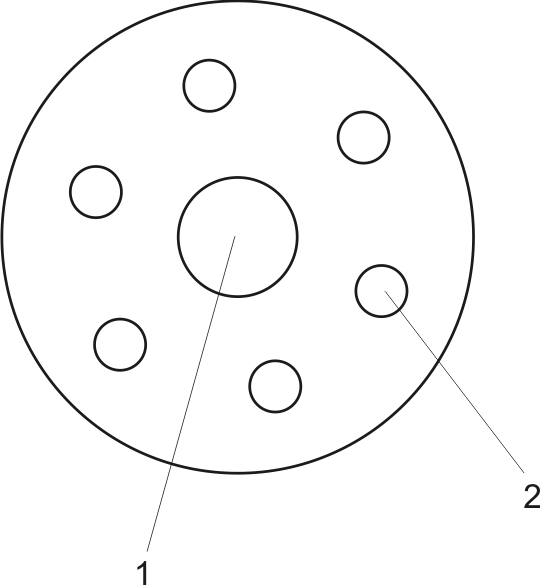
Рис. 7 Поперечный разрез миофиламенты
Между миозином (1) и актиновыми нитями (3) имеются миозиновые поперечные мостики (2). – рисунок 8. В состоянии покоя головка поперечного мостика не соединяется с актином - их соединению препятствует тропомиозин, который находится на глобуле актиновых нитей. Периодически (под влиянием раздражения) тропомиозин двигается в глубину бороздки – в желобок актиновых нитей и освобождает место на актиновой нити для связывания с головкой поперечного мостика.
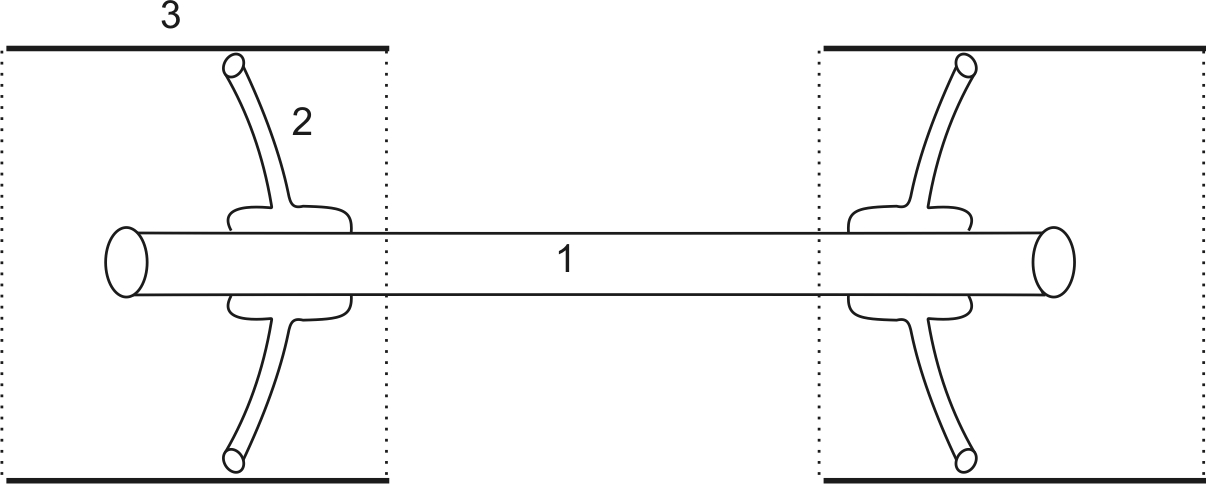
Рис. 8 Соединение миозина с актином
Белки мышц и их роль в мышечном сокращении В механизме мышечного сокращения большую роль играют четыре белка: 1) миозин – этот белок образует толстые нити в структуре миофиламенты, а также участвует в образовании актомиозинового комплекса; 2) актин – этот белок образует тонкие нити в структуре миофиламенты и участвует в образовании актомиозинового комплекса, а при разрыве поперечных мостиков продвигается вдоль миозина, способствуя сокращению мышцы; 3) тропомиозин – это белок регулирует образование актомиозина (обладает аденозинтрифосфотазной активностью), что зависит от локализации тропомиозина: при локализации тропомиозина на глобуле актиновых нитей головка поперечного мостика не соединяется с актином (мышца расслаблена), при локализации тропомиозина в желобке актиновых нитей головка поперечного мостика соединяется с актином и образуется актомиозин, расщепляется АТФ, освобождается энергия нити актина скользят вдоль миозина - происходит сокращение мышцы.; 4) тропонин – этот белок способен изменять свою конформацию (пространственное расположение) в присутствии ионов кальция. Конформационное изменение тропонина, в свою очередь, способствует продвижению тропомиозина в желобок актиновых нитей, благодаря чему головка поперечного мостика соединяется с актином и образуется актомиозиновый комплекс.
Одиночное мышечное сокращение, его фазы. Тетанус и его виды. Одиночное мышечное сокращение – этот вид сокращения возникает на одиночное раздражение (рис.9 а), достаточного для вызова возбуждения мышцы.
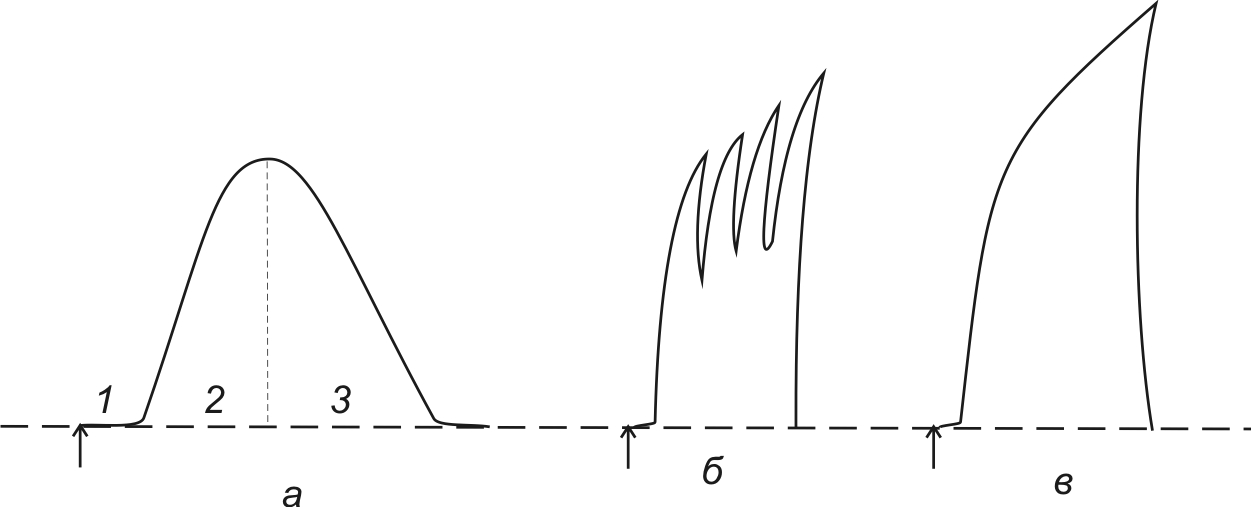
Рис. 9. Одиночное (а) и тетаническое (б, в) сокращение мышц
При этом виде сокращения различают три фазы: 1) латентная фаза (короткий скрытый период) от момента раздражения до появления сокращения; 2) фаза напряжения (сокращения) – в эту фазу происходит нарастание силы сокращения; 3) фаза расслабления – происходит падение силы сокращения до исходной величины. В латентную фазу происходят следующие процессы: деполяризация мембраны - возникновение МПД - распространение пика МПД до Тсистем - выход ионов кальция из цистерн и его соединение с тропонином - изменение конформации тропонина - уход тропомиозина в желобок актиновых нитей - соединение головки поперечного мостика с актином с образованием актомиозинового комплекса -расщепление АТФ. В фазу мышечного сокращение за счет энергии АТФ происходит периодический разрыв поперечного мостика, что приводит к скольжению актиновых нитей вдоль миозина (сокращение мышцы). В фазу расслабления происходят следующие процессы: меньшая часть энергии АТФ идет на работу кальциевого насоса - заход ионов кальция в цистерны - возвращение тропонина к исходной конформации-выход тропомиозина из желобка актиновых нитей-разъединение головки поперечного мостика от актина - возвращение актиновых нитей к исходной позиции (расслабление).
Тетаническое сокращение (рис 9 б, в), или суммированное – при этом происходит длительное и сильное сокращение мышцы. Этот вид сокращения возникает в ответ на ритмический раздражитель – при этом происходит суммация сокращений. Тетаническое сокращение – это сильное и длительное мышечное сокращение, возникающее при действии ритмического раздражителя. В зависимости от частоты ритмического раздражителя различают зубчатый (5-10Гц) и гладкий (более 15 Гц) тетанус. При этом суммация может происходить после частичного расслабления мышцы (зубчатый тетанус – рис. 9 б)) или без частичного расслабления (гладкий тетанус – рис. 9 в).
Продольные и поперечные канальцы (Т-системы). В мышечном волокне имеются множество продольных и поперечных канальцев (рис.10)
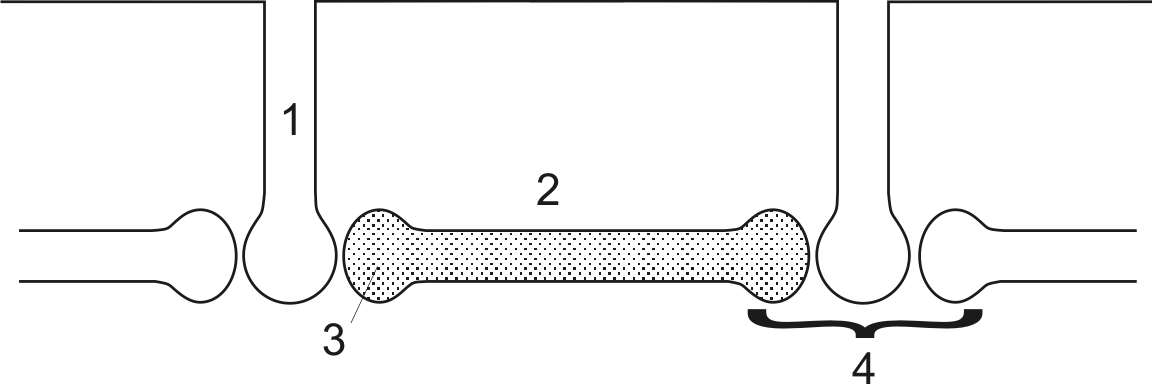
Рис.10. Взаимодействие продольных и поперечных канальцев
Продольные канальцы (2) являются вместилищем ионов кальция (кальциевые цистерны) – в состоянии покоя концентрация ионов кальция в межфибрилярном пространстве очень низкая, так как они находятся в цистернах. Выход ионов кальция из цистерн осуществляется при возникновении МПД, который доходит до этих канальцев (Т-системы мышц). Поперечные канальцы (1) связывают внеклеточную среду с внутриклеточной, а также по мембране этих канальцев осуществляется распространение МПД. Место соединение одного продольного канальца и двух поперечных называю Т-системы мышц.
Последовательность процессов, происходящих при мышечном сокращении. В состоянии покоя головка поперечного мостика не может соединяться с актином, так как этому препятствует тропомиозин, находящийся на актиновой нити. При сокращении мышцы последовательно происходят следующие процессы: раздражение мышцы – возникновение МПД и его распространение по мембране поперечного канальца до Т-систем – увеличивается проницаемость мембраны продольных канальцев (кальциевых цистерн) для ионов кальция – выход кальция в межфибрилярное пространство – изменение конформации тропонина (пространственное распложение молекулы тропонина) – продвижение тропомиозина в желобок актиновых нитей – освобождается место в актиновой нити для соединения головки поперечного мостика с образованием актомиозинового комплекса – гидролиз АТФ с освобождением энергии (все эти процессы происходят в скрытую фазу мышечного сокращения) – большая часть энергии идет на крутящиеся движения поперечных мостиков с последующим их разрывом – скольжение актиновых нитей вдоль миозина, то есть сокращение мышцы. Следует отметить, что каждый разрыв поперечного мостика сопровождается укорочением (напряженим) мышцы. Таким образом, при образовании актомиозинового комплекса происходит расщепление АТФ и выделение энергии. Большая часть энергии используется для того, чтобы создать крутящий момент и отрыв мостика от актиновой нити и благодаря этому актиновая нить продвигается вдоль миозина (скользит) на 10нм. За период укорочения (напряжения) происходит около 50 «гребков», в результате чего длина саркомера уменьшается примерно на 50%.
Последовательность процессов, происходящих при расслаблении мышц. Роль кальцевого насоса в мышечном расслаблении. Меньшая часть энергии, освободившаяся при расщеплении АТФ тратится на работу кальциевого насоса, в результате чего ионы кальция заходят в цистерны и уровень кальция в межфибрилярном пространстве снижается, белок тропонин возвращается к прежней конформации, белок тропомиозин «выходит» из желобка, головка поперчного мостика отталкивается (разъединяется) от актина. Нити актина возвращаются в исходное состояние и наступает процесс расслабления. Таким образом, роль кальциевого насоса заключается в том, что этот насос (энергия) обеспечивает активный транспорт ионов кальция из межфибрилярного пространства (где их мало) в кальциевые цистерны (где их много) и запускает последовательность процессов при расслаблении мышцы.
Современная теория мышечного сокращения. Понятие о состоянии ригор. Процесс сокращения, согласно теории А. Хаксли, Симмонс (1971), происходит за счет скольжения актиновых нитей вдоль миозина за счет крутящегося движения поперечного мостика («гребок») с последующим его разрывом, в результате чего происходит продвижение (скольжение) актиновых нитей вдоль миозина. Скольжения актиновых нитей происходит благодаря целому ряду последовательных процессов: раздражение мышц – возникновение МПД и его распространение по мембране поперечного канальца до Т-систем – увеличивается проницаемость мембраны продольных канальцев (кальциевых цистерн) для ионов кальция – выход кальция в межфибрилярное пространство – изменение конформации тропонина – продвижение тропомиозина в желобок актиновых нитей – соединение головки поперечного мостика с актином и образование актомиозинового комплекса – гидролиз АТФ с освобождением энергии (все эти процессы происходят в скрытую фазу мышечного сокращения) – большая часть энергии идет на крутящиеся движения поперечных мостиков с последующим их разрывом – скольжение актиновых нитей вдоль миозина, то есть сокращение мышцы. Следует отметить, что каждый разрыв поперечного мостика сопровождается укорочением (напряженим) мышцы. Таким образом, при образовании актомиозинового комплекса происходит расщепление АТФ и выделение энергии. Большая часть энергии используется для того, чтобы создать крутящий момент и отрыв мостика от актиновой нити и благодаря этому актиновая нить продвигается вдоль миозина (скользит) на 10нм. За период укорочения (напряжения) происходит около 50 «гребков», в результате чего длина саркомера уменьшается примерно на 50%. В случае отсутствия АТФ наступает стойкое прикрепление головки поперечного мостика к актину без последующего отрыва - такое состояние мышц называется ригор. Это имеет место, например, при трупном окоченении.
Другая часть (меньшая) энергии идет на работу кальциевого насоса – ионы кальция заходят в цистерны – наступает расслабление мышцы.
Функциональная подвижность (лабильность) тканей. Усвоение ритма. Оптимум и пессимум. Способность ткани реагировать на ритмический раздражитель Н.Е. Введенским названа функциональной подвижностью (лабильностью) ткани. Причем, способность ткани воспроизводить циклы возбуждения на ритмический раздражитель зависит от скорости тех элементарных реакций, которыми сопровождается это возбуждение. Поэтому Н.Е. Введенский под лабильностью понимал «большую или меньшую скорость тех элементарных реакций, которыми сопровождается физиологическая деятельность данного аппарата». Из определения следует, что мерой лабильности является максимальное количество циклов возбуждения (пиков МПД), воспроизводимое в единицу времени. Лабильность ткани не является абсолютно постоянной. В ходе ритмического раздражителя лабильность ткани может повышаться – это явление А.А. Ухтомский назвал усвоением ритма. Об усвоении ритма наглядно демонстрирует опыт Мевеса на изолированном нервном волокне лягушки: вначале максимальное количество циклов возбуждений, которое воспроизводило нервное волокно было 460 циклов возбуждений в 1с. При раздражении этого волокна ритмическим раздражителем в 740 импульсов в секунду - нервное волокно воспроизводило каждый второй импульс, но через несколько секунд возникло 740 циклов возбуждений в 1с, то есть нервное волокно усвоило заданный ритм раздражения вследствие повышения лабильности. Н.Е. Введенский в опытах на нервно-мышечном препарате лягушки обнаружил, что если наносить раздражение одной и той же силы, но последовательно увеличивать частоту ритмического раздражителя, то ответная реакция также последовательно нарастает. Им установлено, что с увеличением частоты раздражителя увеличивается ответная реакция. При достижении максимальной величины, ответная реакция постепенно уменьшается и даже прекращается. Частота раздражения (40 – 50гц), вызывающая максимальную ответную реакцию, будет оптимальной, а частота (200 – 400гц), приводящая к уменьшению или исчезновению ответа – пессимальной. Эти реакции объясняются тем, что при оптимальной частоте раздражения каждый последующий импульс попадает в фазу супернормальной возбудимости. При повышении частоты каждый последующий импульс попадает либо в относительную (ответная реакция уменьшается), либо в абсолютную (реакция исчезает полностью) рефрактерность.
Парабиоз и его фазы. Н.Е.Введенский разработал эксперимент с альтерацией (отравлением) нерва на ограниченном участке, тем самым, создав зону пониженной лабильности, то есть возникшее возбуждение в этом участке исчезает медленнее, чем в нормальном (рис. 11)
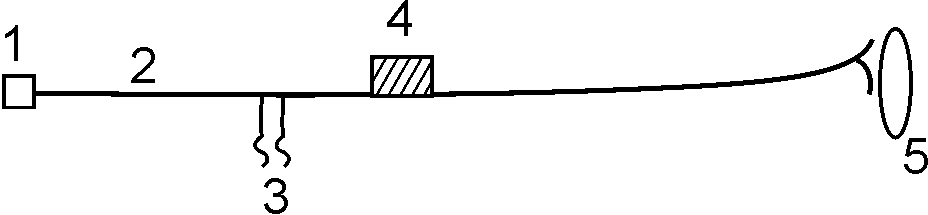
Рис. 11. Опыт Введенского с альтерацией нерва
Альтерацию нерва (2) можно проводить ваткой, смоченной эфиром или хлороформом (4). При этом участок нервного волокна, подвергшийся альтерации, обладает всеми признаками возбуждения (электроотрицательность, изменение физико-химического состояния), кроме одного – этот участок не в состоянии проводить бегущие волны возбуждения. Действуя на электроды (3) ритмическим раздражителем оптимальной частоты, но разной силы (слабые, средние, сильные) Н.Е.Введенский обнаружил несколько последовательно протекающих стадий:
1ст. – трансформирующая, уравнительная, провизорная – при этой стадии на все силы интенсивности раздражения возникает равная по величине реакция; 2ст. – парадоксальная при этом минимальная сила раздражения вызывает более сильный эффект, чем раздражение максимальной силы; 3ст. – тормозная – при этом мышца не реагирует на раздражение любой силы. Тормозная стадия обратимая, но при углублении и усилении альтерации может произойти необратимое нарушение жизнедеятельности – смерть. Именно поэтому обнаруженное явление вследствие альтерации небольшого участка нерва Н.Е. Введенский назвал парабиозом. Парабиоз – состояние близкое к жизни (околожизненное состояние) – активный процесс, который характеризуется неспособностью проводить возбуждение. Наркоз – это частный случай универсальной парабиотической реакции на повреждающее воздействие.
Классификация нервов по структуре. Нерв это совокупность аксонов и дендритов нейрона (нервной клетки). Все нервы по своей структуре делятся на две группы: 1) безмиелиновые нервы, или безмякотные; 2) миелиновые нервы, или мякотные. Безмиелиновые нервы состоят и осевого цилиндра (совокупность отростков) и Швановской оболочки. Миелиновые нервы состоят из: 1) осевого цилиндра; 2) миелина – липид, выполняющий функцию изолятора, а также участвующего в синтезе медиатора; 3) перехваты Ранвье – участок нерва не покрытого миелином; 4) Швановская оболочка. Таким образом, по своей структуре нервы бывают миелиновые и безмиелиновые. Каждый нерв обладает двумя физиологическими свойствами: возбудимостью и проводимостью
Особенности проведения возбуждения по миелиновым и безмиелиновым нервам. Распространение возбуждения по нерву объясняется теорией местных и круговых токов. При действии раздражителя на безмиелиновый нерв происходит деполяризация мембраны в результате чего возникают местные токи и происходит деполяризация мембраны соседних участков слева и справа, возбуждение распространяется в обе стороны (двухстороннее распространение возбуждения). В данном случае возбуждение распространяется по всей поверхности мембраны, поэтому скорость передачи возбуждения в безмиелиновых нервах очень низкая (0,5 – 10 м/с). При действии раздражителя на миелиновый нерв происходит деполяризация мембраны не покрытые миелином (в перехватах Ранвье) в результате чего возникают круговые токи между перехватами Ранвье слева и справа и происходит деполяризация мембраны этих перехватов, возбуждение распространяется в обе стороны (двухстороннее распространение возбуждения). В данном случае возбуждение распространяется от одного перехвата Ранвье к другому или сразу к третьему, поэтому скорость передачи возбуждения в миелиновых нервах очень высокая (до 70 – 120 м/с).
Механизм непрерывного и сальтаторного проведения возбуждения по нервам. Механизм передачи возбуждения по нерву будет зависеть от его структуры. В безмиелиновом нервном волокне происходит непрерывное распространение возбуждения, поэтому местные токи проходят через большую поверхность мембраны. В миелиновых нервах участки, покрытые миелином обладают очень высоким сопротивлением (100000 – 160000 ом/см2). Таким образом, строение миелинового нерва создает предпосылки для качественно нового типа проведения возбуждения – сальтаторного (от латинского persaltum – прыжки). Местные токи, в одном из перехватов не проходят через всю мембрану (как это отмечалось в безмиелиновых нервах), а идут через соседние перехваты, то есть, возникший МПД в участке раздражения, как бы перескакивают («прыгают») от одного перехвата Ранвье к другому. При этом МПД может перескакивать через 2-3 и даже 5 перехватов Ранвье. Нетрудно убедиться в преимуществе сальтаторного типа проведения возбуждения по нерву: 1) при этом увеличивается скорость распространения возбуждения (до 120 м/с); 2) распространение возбуждения происходит с меньшими затратами энергии, то есть более экономно. Скорость проведения импульсов возрастает с увеличением диаметра аксона, так как при этом снижается внутреннее сопротивление.
Законы проведения возбуждения по нервам. Различают три закона проведения возбуждения по нерву. Первый закон свидетельствует о том, что возбуждение распространяется по нерву только в случае его анатомической и физиологической целостности (рис. 12). Так, если перерезать нерв – рис 12б (нарушить анатомическую целостность), то возбуждение по нерву распространяться не будет. Если каким-либо повреждающим агентом (хлороформ, эфир, раствор KCl – рис. 12в) подействовать на нерв, мы нарушаем временно физиологическую целостность нерва. В этом случае возбуждение также не будет распространяться по нерву.
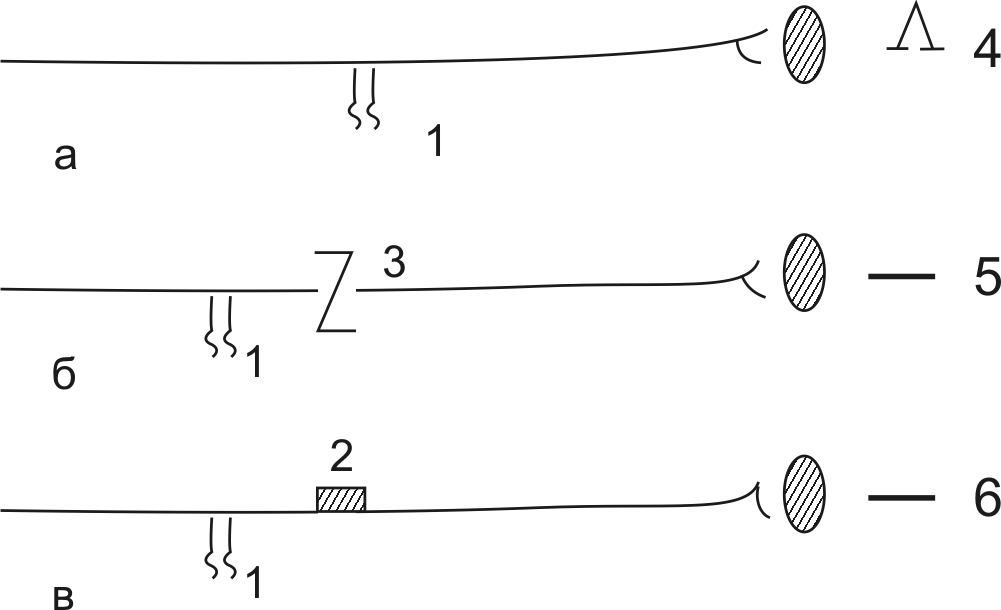
Рис. 12. Закон анатомической и физиологической целостности нерва
Второй закон (рис. 13) свидетельствует о том, что возбуждение распространяется по нерву в обе стороны от места раздражения, то есть в нервах отмечается двухстороннее проведение возбуждения.
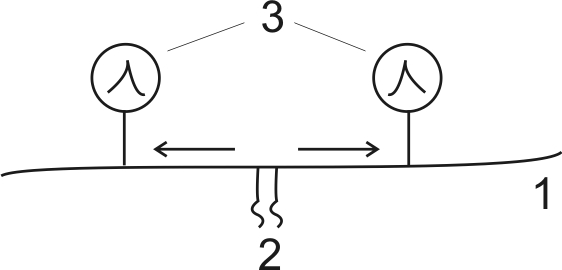
Рис.13. Закон двухстороннего проведения возбуждения
Третий закон (рис. 14) свидетельствует об изолированном проведении возбуждения по нерву, то есть возбуждение с одного нерва (1) не передается на другой нерв (2) как бы близко не находились нервные волокна (участок А-Б).

Рис.14. Закон изолированного проведения возбуждения
Следует отметить, что закон физиологической целостности нерва в какой-то мере относителен для миелиновых волокон, в которых возбуждение распространяется скачкообразно, поэтому при небольшом участке альтерации нерва возбуждение может перейти этот участок. Закон изолированного проведения возбуждения относителен для безмиелиновых нервов, так как при сильном возбуждении оно может перейти с одного безмиелинового нерва на другой безмиелиновый нерв. При местном обезболивании временно нарушается физиологическая целостность нерва, поэтому импульсы от болевых рецепторов не достигают соответствующих зон в коре больших полушарий и человек не чувствует боль.
Синапс, его структура. Нервная система состоит из отдельных нейронов, разделенных межклеточными промежутками, поэтому должен существовать механизм, который позволяет нервному импульсу «перескакивать» с аксона одного нейрона на дендриты или тела другого или, в случае нервно-мышечного соединения - на мышцу.
Синапс – это структурное образование, где происходит переход одного нервного волокна на другой, или переход нерва на нейрон или мышцу (рис. 15)
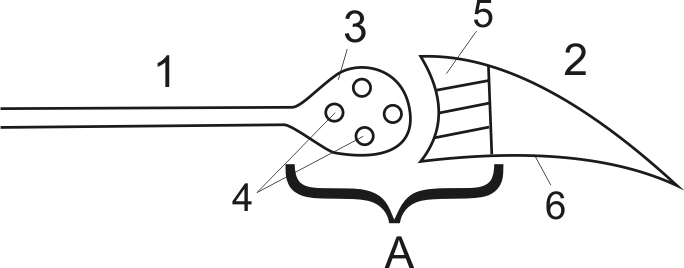
Рис. 15 Структура синапса
Структура любого синапса (А) состоит из: пресинаптической мембраны (3), в которой находятся пузырьки (4), заполненные физиологически активным веществом (медиатор); постсинаптической мембраны (5), которая отличается от внесинаптической мембраны (2) наличием реактивной субстанции, обладающей высокой чувствительностью к медиатору; синаптической щели между пре- и постсинаптической мембраной. Пузырьки пресинаптической мембраны содержат специфическое вещество, которое освобождается при возбуждении аксона и называется медиатором. Окончание аксона с пузырьками называется пресинаптической мембраной. Участок нерва, нейрона или мышцы, куда непосредственно передается возбуждение называется постсинаптической мембраной. Между этими двумя структурами имеется небольшой промежуток (не более 50 нм), который называется синаптической щелью. Из вышеизложенного следует, что в синапсах передача возбуждения осуществляется химическим способом и происходит это за счет трех процессов: 1) освобождения медиатора из пузырьков; 2) диффузии медиатора в синаптическую щель; 3) соединением этого медиатора со специфическими реактивными структурами постсинаптической мембраны, что приводит к образованию нового импульса.
Механизм передачи возбуждения через синапс. ВПСП и ТПСП. При каждом нервном импульсе происходит освобождение медиатора из пузырьков, который диффундирует в синаптическую щель и соединяется с реактивными структурами (субстанциями, рецепторами) постсинаптической мембраны и при этом возникает усиление проницаемости для ионов натрия (возбуждающие медиаторы) или ионов калия (тормозные медиаторы).
За счёт возбуждающего медиатора увеличивается проницаемость для ионов натрия и происходит деполяризация постсинаптической мембраны с образованием возбуждающего постсинаптического потенциала (ВПСП). Если ВПСП не доходит до критического уровня, возникает локальный ответ. Если ВПСП доходит до критического уровня (это зависит от количества медиатора), тогда во внесинаптической мембране возникает МПД. За счет тормозоного медиатора увеличивается проницаемость для ионов калия и происходит гиперполяризация мембраны с образованием тормозного постсинаптического потенциала (ТПСП). Возникновение ВПСП или ТПСП на постсинаптической мембране зависит от медиатора, который выделяется в пресинаптической мембране и от реактивной субстанции в постсинаптической мембране. Например, ацетилхолин может вызвать образование ВПСП и ТПСП: если ацетилхолин взаимодействует с Н-холинореактивной субстанцией повышается проницаемость постсинаптической мембраны для ионов натрия и возникает ВПСП, если ацетилхолин взаимодействует с М-холинореактивной субстанцией повышается проницаемость для ионв калия и возникает ТПСП.
Холинореактивные и адренореактивные субстанции. В постсинаптической мембране синапса может быть два вида белков чувствительных к медиатору (эти белки называются реактивной субстанцией): 1) холинореактивные субстанции, обладающие высокой чувствительностью к медиатору ацетилхолину. В зависимости от повышения проницаемости постсинаптической мембраны для ионов калия или натрия холинореактивные субстанции делятся на а) Н-холинореактивные – при взаимодействии их с ацетилхолином увеличивается проницаемость для ионов натрия и на постсинаптической мембране будет деполяризация, возникает возбуждающий постсинаптический потенциал (ВПСП); б) М-холинореактивные субстанции - при взаимодействии их с ацетилхолином увеличивается проницаемость для ионов калия и на постсинаптической мембране будет гиперполяризация, возникает тормозной постсинаптический потенциал (ТПСП); 2) адренореактивные субстанции, обладающие высокой чувствительностью к медиатору норадреналину. В зависимости от повышения проницаемости постсинаптической мембраны для ионов калия или натрия адренореактивные субстанции делятся на а) альфа, и бета1 адренореакивные субстанции – при взаимодействии их с норадреналином увеличивается проницаемость для ионов натрия и на постсинаптической мембране будет деполяризация, возникает ВПСП; б) бета2 адренореактивные субстанции - при взаимодействии их с норадреналином увеличивается проницаемость для ионов калия и на постсинаптической мембране будет гиперполяризация, возникает ТПСП.
АНС, его отделы и функции. По ныне действующей анатомической номенклатуре все термины (вегетативная нервная система, висцеральная, чревная, непроизвольная) заменены на «автономную нервную систему» (АНС). Однако в нашей литературе пользуются термином «вегетативная нервная система» (ВНС). Той же анатомической номенклатурой АНС делится на симпатический и парасимпатический отделы.
АНС включает следующие образования: 1) нервные волокна (пре- и постганглионарные), 2) переферические нервные узлы (ганглии), состоящие из нейронов, 3) центров в сером веществе головного (мезенцефальной и бульбарной области) и спинного мозга (боковых рогах), 4) высших центров, находящихся в межуточном мозге (гипоталамусе). АНС состоит из двух отделов: симпатического и парасимпатического.
Функции АНС: 1) обеспечивает местное и общее ауторегулирование (саморегуляцию) органов и систем организма в целях сохранения относительного динамического постоянства внутренней среды (гомеостазиса); 2) обеспечивает адаптацию вегетативных функций к изменяющимся условиям внешней и внутренней среды; 3) обеспечивает трофическую функцию органов и тканей. Трофическое влияние заключается в регулирующем влиянии АНС на обмен веществ и питание органов и тканей.
Локализация нейронов симпатического и парасимпатического отделов. Нейроны симпатического и парасимпатического отделов АНС локализуются в центральной части, которую можно разделить на 4 области: 1) мезенцефальная (средний мозг), здесь находятся нейроны, отростки которых образуют парасимпатические нервы I-IV пары черепно-мозговых нервов; 2) бульбарная (продолговатый мозг) – здесь находятся нейроны, отростки которых образуют восемь пар черепно-мозговых нервов (V – ХII пары), также относящихся к парасимпатическим нервам; 3) сакральная (в боковых рогах II – IV крестцовых сегментов спинного мозга) – осуществляется парасимпатическая иннервация мочеполовых органов и нижних отделов пищеварительного тракта. 4) тораколюмбальная (боковые рога грудных и поясничных сегментов спинного мозга), здесь находятся нейроны, отростки которых образуют симпатические нервы, которые осуществляют иннервацию всех органов и тканей организма.
Отличия АНС и соматической нервной системы. Основные отличия АНС от цереброспинальной (соматической) нервной системы:
1. По расположению двигательных нейронов: для соматической нервной системы двигательные нейроны (мотонейроны) расположены в сером веществе спинного мозга (в передних рогах). Для АНС первый нейрон расположен в мезенцефальной и бульбарной области головного мозга (парасимпатический отдел АНС) а также в боковых рогах спинного мозга (симпатический – в шейных, грудных и поясничных сегментах, парасимпатический – в крестцовых сегментах). Вторые нейроны рассеяны на периферии: а) скопление нейронов в позвоночных ганглиях, находящихся на телах позвонков в составе пограничного симпатического ствола; б) скопление нейронов в предпозвоночных узлах (солнечное сплетение, нижнее брыжеечное сплетение, сердечное сплетение и т.д.); в) парасимпатические ганглии залегают внутри органа – интрамуральные ганглии.
2. По конечному результату: эфферентные волокна соматической нервной системы заканчиваются в скелетных мышцах, а эфферентное волокно АНС – во всех внутренних органах и гладкой мускулатуре сосудов.
3. По связи эффектора (рабочего органа) с ЦНС: для соматической нервной системы эта связь однонейронная, то есть аксон мотонейрона, находящегося в передних рогах спинного мозга, заканчивается в скелетных мышцах. Для АНС – эта связь двухнейронная, поэтому эфферентное нервное волокно АНС состоит из двух частей: преганглионарного волокна (его нейрон находится в спинном и головном мозгу) и постганглионарного (его нейрон находится в переферическом ганглии).
4. Выходом нервных волокон: волокна соматической нервной системы выходят из ЦНС строго сегментарно; волокна АНС подразделяются на симпатические и парасимпатические. Симпатические волокна выходят из центров, расположенных в боковых рогах шейных, грудных и поясничных сегментов. Парасимпатические волокна выходят из центров, расположенных в среднем и продолговатом мозге, а также в боковых рогах спинного мозга крестцовых сегментов.
5. По структуре нервного волокна: соматические нервные волокна в основном относятся к типу А – миелиновые волокна большого диаметра (в среднем 15 мкм.) и большой скоростью проведения возбуждения (в среднем 100 м/с). Преганглионарные волокна АНС в большей части относятся к типу В – миелиновые волокна меньшего диаметра (в среднем 3 мкм.) и меньшей скоростью (в среднем 7 м/с) Постганглионарные волокна АНС в большей части относятся к волокнам типа С – немиелиновые, маленького диаметра (в среднем 1 мкм) и маленькой скоростью проведения возбуждения (в среднем 1 м/с).
6. По медиатру: в окончаниях соматического нерва выделяется только ацетилхолин; в окончаниях симпатического нерва выделяется ацетилхолин (в окончаниях преганглионарного волокна) и норадреналин (в окончаниях постганглионарного волокна); в окончаниях пре- и постганглионарного волокна парсимпатического нерва выделяется только ацетилхолин.
7. По реактивной субстанции в постсинаптической мембране (ПСМ): в ПСМ соматического синапса отмечается только Н-холинореактивная субстанция; в ПСМ симпатического и парасимпатического ганглия отмечается Н-холинореактивная субстанция; в ПСМ рабочего органа симпатического нерва может быть либо альфа, либо бета1, либо бета2 адренореактивная субстанция; в ПСМ рабочего органа парасимпатического нерва может быть либо М-, либо Н-холинореактивная субстанция.
Отличия симпатического и парасимпатического отделов АНС. Основные отличия симпатического и парасимпатического отделов АНС:
1. По расположению двигательных нейронов: для парасимпатического отдела АНС первый нейрон расположен в мезенцефальной (I-IV пары черпномозговых нервов) и бульбарной (V-XII пары черепномозговых нервов) области головного мозга, а также в боковых рогах спинного мозга (в крестцовых сегментах). Второй нейрон залегает внутри органа – интрамуральные ганглии; для симпатического отдела АНС первый нейрон расположен в боковых рогах спинного мозга шейных, грудных и поясничных сегментах. Второй нейрон расположен в виде: а) скопление нейронов в позвоночных ганглиях, находящихся на телах позвонков в составе пограничного симпатического ствола (паравертебральные ганглии); б) скопление нейронов в предпозвоночных узлах (солнечное сплетение, нижнее брыжеечное сплетение, сердечное сплетение и т.д.).
2. По эфферентному волокну: в симпатическом нерве преганглионарное волокно короче постганглионарного, а в парсимпатическом нерве – наоборот (преганглионарное волокно длинее постганглионарного)
3. По медиатру: в окончаниях симпатического нерва выделяется ацетилхолин (в окончаниях преганглионарного волокна) и норадреналин (в окончаниях постганглионарного волокна); в окончаниях пре- и постганглионарного волокна парсимпатического нерва выделяется только ацетилхолин.
4. По реактивной субстанции в постсинаптической мембране (ПСМ): в ПСМ рабочего органа симпатического нерва может быть либо альфа, либо бета1, либо бета2 адренореактивная субстанция; в ПСМ рабочего органа парасимпатического нерва может быть либо М-, либо Н-холинореактивная субстанция. В ПСМ симпатических и парасимпатических ганглиев находится только Н-холинореактивная субстанция
Медиаторы АНС и СНС. Эфферентные нервы соматического нерва однонейронный и в его окончаниях выделяется медиатор ацетилхолин.Эфферентные нервы АНС двухнейронные, поэтому здесь дважды выделяется медиатор: в синапсах между пре- и постганглионарными волокнами (в ганглиях) и между постганглионарным волокном и органом. Симпатический нерв по выделяемому медиатору не однороден: в окончаниях преганглионарных волокон выделяется ацетилхолин, а в окончаниях постганглионарных волокон выделяется норадреналин. Некоторые симпатические постганглионарные нервные волокна выделяют ацетилхолин (потовые железы). Парасимпатический нерв по выделяемому медиатору однороден, так как в пре- и постганглионарных волокнах выделяется ацетилхолин. В органах, иннервируемых АНС имеются специальные белковые структуры, обладающие очень высокой чувствительностью к медиатору. Выше было отмечено, что в постганглионарных волокнах симпатического нерва выделяется норадреналин, а парасимпатического нерва – ацетилхолин, поэтому все белковые структуры органов можно разделить на две группы:
I.Белковые структуры, обладающие высокой чувствии-тельностью к норадреналину – это адренореактивные структуры. Было отмечено, что раздражение симпатического нерва вызывает разную реакцию различных органов. Например, при раздражении симпатического нерва сердца происходит увеличение всех свойств сердечной мышцы и работа сердца усиливается, а при раздражении симпатического нерва кишечника – уменьшается тонус мышц кишечника. Это различие связано с различными адренореактивными структурами, имеющимся в этих органах. Различают три вида адренореактивных структур: 1) альфа адренореактивные структуры – при взаимодействии этих структур с норадреналином происходит усиление функции; 2) бета1 – адренореактивные структуры – при взаимодействии этих структур с норадреналином также происходит усиление функции органа; 3) бета2 – адренореактивные структуры – при взаимодействии этих структур с норадреналином происходит ослабление функции органа. Таким образом, при раздражении симпатического нерва результат работы органа будет разным в зависимости от того, какие адренореактивные структуры превалируют в этом органе.
II.Белковые вещества, обладающие высокой чувствительностью к ацетилхолину – холинореактивные структуры. В свою очередь, раздражение парасимпа-тического нерва также может привести к ослаблению или усилению функции, что зависит от различных видов холинореактивных структур: 1) М-холинореактивные структуры, котрые чувствительны к ацетилхолину и мускарину (яду грибов). При взаимодействии ацетилхолина с этими структурами происходит ослабление функции органа; 2) Н-холинореактивные структуры – они чувствительны к ацетилхолину и никотину. При взаимодействии ацетилхолина с этими структурами происходит усиление функции органа. Таким образом, конечный результат функции органа при раздражении парсимпатического нерва зависит от преимущественного содержания в этом органе М- или Н-хлинореактивных структур. Так, в сердечной мышце превалируют М-холинореактивные структуры, а в мышечном слое кишечника Н-холинореактивные структуры, поэтому при раздражении парасимпатического нерва сила сокращения сердечной мышцы уменьшается или мышца не сокращается (остановка сердца), а сила сокращения мышц кишечника увеличивается (усиливается моторика кишечника). Следует отметить, что в постсинаптической мембране ганглиев и скелетных мышц имеются только Н-холинореактивные структуры.
Синергизм и антагонизм отделов АНС. Типы взаимодействия отделов АНС, способ определения. По основным эффектам при раздражении холинергических и адренергических нервов можно выделить два типа взаимоотношения симпатического и парасимпатического отделов АНС: 1) чаще всего отмечается антагонизм. Так, сердечная мышца при раздражении симпатического нерва усиливает свое сокращение, а при раздражении парасимпатического нерва – уменьшается сила сокращения; гладкая мускулатура бронхов и кишечника при раздражении симпатического нерва расслабляется, а при раздражении парасимпатического нерва – усиливает свое сокращение; 2) в ряде случаев отмечается синергизм – однонаправленное действие симпатического и парасимпатического нервов. Так, при раздражении симпатического нерва усиливается сокращение радиарных мышц зрачка (зрачок расширяется), а при раздражении парасимпатического нерва тоже усиливается сокращение, но уже циркулярных мышц зрачка (зрачок суживается). По конечному результату вроде бы отмечается антагонизм (в одном случае зрачок расширяется, в другом – суживается), но по влиянию непосредственно на мышцы – отмечается синергизм в обоих случаях усиливается сокращение мышц. Синергизм можно отметить и при действии этих нервов на слюнные железы: при раздражении парасимпатического нерва усиливается слюноотделение жидкой консистенции, а при раздражении симпатического нерва количество слюны либо не меняется, либо несколько увеличивается, но при этом слюна становится вязкой (трофическое влияние симпатического нерва). Существуют органы, снабжаемые только симпатическими нервами (почти все кровеносные сосуды, селезенка, гладкие мышцы волосяных луковиц) или только парасимпатическими (циркулярные мышцы зрачка, слезная железа) нервами. Под действием симпатических нервов может усиливаться глюкогенолиз в печени и липолиз в жировых клетках, что приводит к увеличению концентрации глюкозы и свободных жирных кислот в крови. Парасимпатические нервы не влияют на эти процессы.
Под влиянием симпатического и парасимпатического отдела АНС на орган можно выделить три типа взаимодействия:
1) нормотонический тип – при этом отмечается оптимальная координация симпатического и парасимпатичес-кого отделов АНС в регуляции работы органа;
2) симпатикотонический тип – при этом отмечается дискоординация во влиянии отделов АНС на орган с преимущественным влиянием симпатического нерва;
3)ваготонический тип – при этом отмечается также дискоординация во влиянии отделов АНС на орган, но с преимущественным влиянием парасимпатического нерва. Типы взаимодействия отделов АНС можно определить при помощи корреляционных ритмограмм.
Особенности постсинаптической мембраны в ганглиях и эффекторах. В постсинаптической мембране симпатического и парасимпатического ганглия находятся только Н-холинореактивные субстанции, которые соединяются с ацетилхолином и возникает возбуждающий постсинаптический потенциал за счет повышения проницаемости для ионов натрия. Благодаря возникновению ВПСП импульс передается от преганглионарного волокна на постганглионарное и дальше к рабочему органу.
В окончаниях постганглионарного волокна парасимпатического нерва выделяется ацетилхолин, поэтому в постсинаптической мембране эффектора могут быть Н- и М-холинореактивные структуры. При взаимодействии ацетилхолина с Н-холинореактивной структурой увеличивается проницаемость для инов натрия и возникает ВПСП – усиливается функция эффектора. При взаимодействии ацетилхолина с М-холинореактивной структурой увеличивается проницаемость для инов калия и возникает ТПСП – уменьшается функция эффектора.
В окончаниях постганглионарного волокна симпатического нерва выделяется норадреналин, поэтому в постсинаптической мембране эффектора могут быть альфа, бета1 или бета2 адренореактивные структуры. При взаимодействии норадреналина альфа и бета1 адренореактвной структурой увеличивается проницаемость для ионов натрия и возникает ВПСП – усиливается функция эффектора. При взаимодействии норадреналина с бета2 адренореактивной структурой увеличивается проницаемость для инов калия и возникает ТПСП – уменьшается функция эффектора.
Нейрон, как структурная и функциональная единица ЦНС, их классификация. Структурно-функциональной единицей ЦНС является нейрон (нервная клетка). Он состоит из тела (сомы) и отростков – многочисленных дендритов и одного аксона. Дендриты (короткие отростки) сильно ветвятся и образуют множество синапсов с другими нейронами, что определяет их ведущую роль в восприятии нейроном информации. Аксон начинается от тела аксонным холмиком, функцией которого является генерация нервного импульса, который по аксону проводится к другим клеткам. Аксон сильно ветвится, образуя множество коллатералей, терминали которых образуют синапсы с другими клетками.Многочисленные нейроны можно разделить на следующие:
Афферентные нейроны, или чувствительные, они самые многочисленные и локализуются в задних рогах спинного мозга. Аксоны этих нейронов заканчиваются на различных рецепторах и фактически составляют афферентные пути, то есть пути, по которым сигналы поступают в ЦНС.
Мотонейроны, или двигательные нейроны. Различают три вида основных мотонейронов:
а) альфа-мотонейроны – они локализуются в передних рогах спинного мозга, их аксоны заканчиваются на экстрафузальных мышцах;
б) гамма-мотонейроны – также локализуются в передних рогах спинного мозга, их аксоны заканчиваются в интрафузальных мышцах;
в) вегетативные мотонейроны - локализуются в боковых рогах спинного мозга, их аксоны заканчиваются на внутренних органах и сосудах и составляют симпатические и парасимпатические нервы. Из вышеизложенного следует, что аксоны всех мотонейронов образуют эфферентные пути, то есть пути, по которым сигналы из ЦНС поступают в рабочие органы.
Гигантские тормозные клетки Реншоу. Отростки этих нейронов заканчиватся на альфа мотонейронах, в их окончаниях выделяется тормозной медиатор гамма аминомасляная кислота (ГАМК), который вызывает их торможение за счет гиперполяризации. При возбуждение этих нейронов происходит процесс торможения в альфа мотонейронах и возникает торможение спиномозговых рефлексов.
Тормозные интернейроны – их отростки заканчиватся на клетках Реншоу. При возбуждении тормозных интернейронов происходит торможение клеток Реншоу и возникает облегчение спиномозговых рефлексов.
Особенности проведения возбуждения в ЦНС (замедление скорости проведения возбуждения, одностороннее проведение возбуждения, конвергенция, дивергенция, иррадиация). Все особенности проведения возбуждения в ЦНС связаны с наличием большого количества синапсов:
1) Замедление проведения возбуждения по ЦНС. Эта особенность обусловлена тем, что в ЦНС имеются много синапсов, а в каждом синапсе передача возбуждения осуществляется за счет медиатора – необходимо время для выделения медиатора, его диффузии через пресинаптическую мембрану и взаимодействие с реактивной субстанцией постсинаптической мембраны с возникновением ВПСП.
2) Одностороннее проведение возбуждения, так как в синапсах возбуждение передается химическим способом – с помощью медиатора, который выделяется в пресинаптической мембране, поэтому возбуждение может передаваться только от пресинаптической мембраны к постсинаптической мембране.
3) Конвергенция, то есть схождение импульсов к одному нейрону. Эта особенность обусловлена тем, что рецепторов значительно больше, чем нейронов, поэтому от нескольких рецепторов (одного и того же рецептивного поля) импульсы поступают к одному нейрону – это и есть конвергенция.
4) Дивергенция - иррадиация – расхождение импульсов в ЦНС от одного нейрона к другому. Расхождение импульсов в ЦНС обусловлено наличием большого количества вставочных нейронов и многочисленных отростков. При сильном раздражении рецептора происходит распространение (иррадиация) возбуждения в ЦНС от одного нейрона к другим и даже к тем нейронам, которые не относятся к данному рецептивному полю. В результате иррадиации процессом возбуждения охватываются большое количество эффекторов (рабочих органов), и даже тех, которые не относятся к данному рецептивному полю. Из выше изложенного следует, что иррадиация является следствием дивергенции.
Пространственная и последовательная суммация возбуждения в ЦНС. Различают два вида суммаций:
1) пространственная, или одновременная. Этот вид суммации возникает благодаря тому, что рецепторов значительно больше, чем афферентных нейронов, поэтому из 2-х и более рецепторов сигналы поступают в один и тот же нейрон. Для того, чтобы убедиться в наличии данной суммации, необходимо подействовать на рецептор подпороговой силой – никакого ответа не будет. Затем подействовать одновременно двумя подпороговыми силами на два рецептора одного и того же рецептивного поля (совокупность рецепторов при раздражении которых происходит одна и та же реакция) – будет ответная реакция. При действии одной подпороговой силой выделяется мало медиатора и ВПСП на постсинаптической мембране не доходит до критического уровня, поэтому нет ответной реакции. При одновременном действии двух подпороговых сил медиатор выделяется одновременно в двух нервных окончаниях, которые заканчиваются на одном и том же нейроне. Таким образом, благодаря суммации медиатора в синаптической щели его становится достаточно для того, чтобы ВПСП дошел до критического уровня и возник МПД во внесинаптической мембране;
2)последовательная суммация, или временная. Этот вид суммации происходит при действии ритмического раздражителя. При этом частота ритмического раздражителя должна быть достаточно высокой, чтобы медиатор, выделившийся на первый стимул не успел разрушиться и выделяется новая порция медиатора от последующих раздражений. В данном случае медиатор уммируется в пресинаптической мембране. Из выше изложенного следует, что при обоих видах суммаций происходит суммация медиатора. При пространственной (одновременной) суммации медиатор суммируется в синоптической щели. При последовательной (временной) суммации медиатор суммируется в пресинаптической мембране.
Последействие в ЦНС. Различают два вида последействия:
а) кратковременное последействие – его причиной является высокий уровень следовой деполяризации в нейронах ЦНС. Кратковременное последействие может возникнуть при действии порогового раздражителя. Это связано с особенностью возникновения МПД в нейронах и с низким порогом раздражения: разница между МПП (-70 мв) и критическим уровнем деполяризации (-60 мв). При возникновении МПД в нейронах возникает высокий уровень следовой деполяризации, поэтому одиночное раздражение вызывает несколько реакций: первая реакция в ответ на раздражение, остальные (последействие) за счет высокого уровня следовой деполяризации.
б) длительное последействие – его причиной является циркуляция импульсов в ЦНС по коллатералям через вставочные нейроны. Длительное последействие возникает на действие максимального или сверхмаксимального раздражителя. При этом происходит длительная циркуляция импульса в ЦНС, через вставочный возбуждающий нейрон, поэтому при одиночном раздражении рецептора в ЦНС длительно сохраняется возбуждение и отмечается длительная ответная реакция эффектора после прекращения раздражения рецептора.
Окклюзия. Окклюзия, закупорка – уменьшение величины ответной реакции при одновременном раздражении двух и более рецепторов по сравнению с суммарной реакцией, полученной при раздражении каждого рецептора в отдельности (рис. 16).

Рис. 16. Механизм возникновения окклюзии
При раздражении каждого из двух рецепторов (А и Б) в отдельности в ЦНС происходит возбуждение по 4 нейрона (при раздражении рецептора А возбуждаются нейроны 1,2,3,4; при раздражении рецептора Б также возбуждаются 4 нейрона 3,4,5,6). Причем, два нейрона (3 и 4) являются общими для каждого рецептора, то есть эти нейроны возбуждаются при раздражении каждого рецептора в отдельности. При одновременном раздражении обеих рецепторов в ЦНС возбуждаются не 8 нейронов, а 6, так как два нейрона являются общими. Эти два нейрона возбуждаются по конкурентному механизму либо за счет раздражения рецептора А, либо за счет раздражения рецептора Б. Если эти нейроны возбуждаются за счет импульсов, возникающих при раздражении одного рецептора, то определенное время эти нейроны будут находиться в рефрактерной фазе возбудимости и не реагируют на импульсы, возникающие при раздражении другого рецептора. Таким образом, в результате окклюзии (закупорки импульсов, возникающих при раздражении обеих рецепторов) при одновременном раздражении обеих рецепторов реакция обеих эффекторов будет меньше суммы реакции каждого эффектора при раздражении рецепторов в отдельности. Из выше изложенного видно, что окклюзия является следствием конвергенции.
Виды торможения в ЦНС. Впервые о торможение в ЦНС высказал И.М.Сеченов (1861 год). Исследуя рефлекторную деятельность лягушки с сохраненными зрительными буграми. И.М.Сеченов определял время сгибательного рефлекса – в ответ на погружение лапки лягушки в кислоту происходило сгибание конечности в тазобедренном и коленном суставах. При помещении на зрительные бугры кристаллы поваренной соли, возникает торможение, о чем свидетельствует удлинение времени рефлекса. Это наблюдение и позволило И.М. Сеченову говорить о явлении торможения в ЦНС. В последующем такой вид торможения получил название сеченовское торможение, или центральное торможение.
По своей локализации различают два вида торможения в ЦНС: 1) пресинаптическое, которое локализовано в пресинаптических терминалях, за счет постоянного возбуждения возбуждающих вставочных нейронов; 2) постсинаптическое – локализовано в постсинаптической мембране. К этому виду торможения относятся: сеченовское, реципрокное (сопряженное), возвратное и пессимальное.
Виды постсинаптического торможения, их механизм. Постсинаптическое торможение локализуется в постсинаптической мембране. Различают следующие виды:
1) Сеченовское торможение: после препарирования зрительных бугров у лягушки проверяем время рефлекса. Для этого наносим раздражение (раствором серной кислоты) на рецептор и возникает ответная реакция (отдергивание лапки – рефлекс). Определяем время рефлекса (время от момента раздражения до появления ответной реакции). После этого наносим кристаллики поваренной соли на зрительные бугры. В результате при действии прежнего раздражителя на рецептор требуется больше времени для возникновения ответной реакции – происходит торможение;
2) Реципрокное торможение обеспечивает осуществлению противоположных функций (вдоха и выдоха, сгибание и разгибание, отведение и приведение и т.д.). Реципрокное торможение можно рассмотреть на примере коленного рефлекса – разгибание голени. В данном случае удар неврологическим молоточком по сухожилию мышц разгибателей голени происходит растяжение этой мышцы, возбуждаются рецепторы мышечного веретена. Импульсы от этого рецептора через афферентные нейроны одновременно поступают в тормозные клетки Реншоу и альфа мотонейроны мышц разгибателей голени. При возбуждении клеток Реншоу происходит торможение альфа мотонейронов мышц сгибателей голени за счет гиперполяризации. Таким образом, при ударе неврологическим молоточком по сухожилию мышц разгибателей голени одновременно происходит возбуждение альфа мотонейронов мышц разгибателей голени и торможение альфа мотонейронов мышц сгибателей – отмечается разгибание голени (коленный рефлекс). Такое торможение нейронов, которое происходит при одновременном возбуждении другого нейрона получило название реципрокного, или сопряженного, так как этот вид торможения связан (сопряжен) с возбуждением другого нейрона;
3) Возвратное торможение. При сильном раздражении рецептора импульсы идут не только к эффектору, но по коллатерали возвращаются к нейрону через клетки Реншоу, вызывая его гиперполяризацию (торможение). Таким образом, альфа-мотонейрон, активируясь, через систему тормозного нейрона сам себя тормозит, за счет возвратных импульсов;
4) Пессимальное торможение возникает при действии ритмического раздражителя высокой частоты. В основе механизма сеченовского, реципрокного и возвратного торможения лежит гиперполяризация постсинаптической мембраны, которая происходит при возбуждении клеток Реншоу. В основе механизма пессимального торможения лежит стойкая деполяризация постсинаптической мембраны, в результате чего блокируются натриевые каналы и происходит торможение.
Доминантный очаг возбуждения, его свойства. Принцип доминанты - был открыт А.А. Ухтомским. Изучая ответы скелетной мышцы кошки на электрические раздражения коры больших полушарий, он обнаружил, что при акте дефикации ответы мышцы прекращаются. Проанализировав этот факт А.А. Ухтомский пришел к мнению о наличии в ЦНС явления доминанты – господствующего очага возбуждения. Дело в том, что среди многочисленных рефлекторных актов, которые могут быть выполнены в данный момент, осуществляются те реакции, которые наиболее значимы для организма в данный момент времени. Центры, участвующие в реализации более значимых для организма реакций, А.А. Ухтомский назвал «доминантным очагом возбуждения». Этот «очаг» обладает рядом важных свойств: а) повышенной возбудимостью; б) стойкостью возбуждения (инертностью), поэтому этот очаг возбуждения трудно подавить другим очагом возбуждения; в) способностью к суммации субдоминантных возбуждений, то есть доминантный очаг возбуждения притягивает к себе другие очаги возбуждения, менее значимые для организма в данный момент времени; г) способностью тормозить субдоминантные очаги возбуждения.
Обратная связь, его виды. ЦНС посылает к рабочим органам определенные команды (каким должен быть результат). Фактический результат работы органа несколько отличается от должного. Разница фактического и должного результата называется рассогласованием: чем больше рассогласование, тем менее оптимальными являются процессы управления (регулирования). Обратная связь несет информацию в ЦНС о параметрах фактического результата и в ЦНС осуществляется сравнение фактических параметров с теми, которые должны быть в данных условиях. Если фактический результат больше заданного уменьшается функция эффектора (отрицательная обратная связь), если фактический результат меньше заданного – усиливается функция эффектора (положительная обратная связь). Из вышеизложенного следует, что благодаря сигналам по обратной связи происходит коррекция (исправление) командных сигналов от ЦНС, чтобы приблизить параметры фактического результата к должным параметрам – в результате уменьшается рассогласование и происходит оптимизация процессов управления (регуляции). При положительной обратной связи происходит усиление параметров результата, а при отрицательной - уменьшение.
Функции спинного мозга (фазические и тонические рефлексы). Спинной мозг расположен в спиномозговом канале и выполняет две основные функции: 1) рефлекторную и 2) проводниковую. Все рефлексы спинного мозга можно разделить на: 1) соматические (тонические и фазические) и 2) вегетативные. В 1811 году Белла, разрушая передние корешки спинного мозга поясничных сегментов, отметил потерю двигательной функции задних конечностей. В 1822 году Мажанди, разрушая задние корешки спинного мозга поясничных сегментов, отметил потерю чувствительности кожи нижних конечностей. Таким образом, в задних корешках проходят чувствительные центростремительные нервные волокна от рецепторов кожи (болевые, температурные, тактильные и давления), мышц,сухожилий, суставов (проприоцептивная система) и от рецепторов внутренних органов (вегетативная система); в передних корешках проходят двигаьтельные центробежные нервные волокна от нейронов к мышцам. Эта особенность функции задних и передних корешков спинного мозга отражается законом Белла-Мажанди.
В соматических рефлексах участвуют альфа и гамма мотонейроны в передних рогах спинного мозга. Аксоны этих нейронов заканчиваются на экстрафузальных (альфа мотонейроны) и интрафузальных (гамма мотонейроны) мышцах. К фазическим относится все антогонистические рефлексы (сгибание-разгибание, отведение-приведение и т.д.). Эти рефлексы осуществляются фазно: мышцы быстро сокращаются и быстро расслабляются. Как правило, в этих рефлексах участвуют белые мышцы (мышцы I типа). Тонические рефлексы – это длительное сокращение мышц. В этих рефлексах участвуют красные мышцы (IIA и IIB типы). Тонические рефлексы можно разделить на периферические и центральные. Периферические тонические рефлексы осуществляются при расслаблении экстрафузальных мышц, что приводит к растяжению ядерной сумки мышечного веретена (рецепторов растяжения скелетных мышц). При этом происходит возбуждение рецепторов ядерной сумки, импульсы по афферентным волокнам через задние корешки спинного мозга поступают в альфа мотонейроны передних корешков и отсюда по эфферентным волокнам импульсы идут в экстрафузальные мышцы, происходит их сокращение. Центральные тонические рефлексы осуществляются за счет ретикуло-спинального пути. Импульсы от нейронов РФ мозга поступают в гамма мотонейроны передних корешков спинного мозга, отсюда по эфферентным путям к интрафузальным мышцам, при сокращении которых происходит натяжение ядерной сумки. При этом происходит возбуждение рецепторов ядерной сумки, импульсы по афферентным волокнам через задние корешки спинного мозга поступают в альфа мотонейроны передних корешков и отсюда по эфферентным волокнам импульсы идут в экстрафузальные мышцы, происходит их сокращение. Таким образом, натяжение ядерной сумки мышечного веретена (возбуждение проприорецепторов мышц) происходит двумя способами: 1) за счет расслабления экстрафузальных мышц и 2) за счет сокращения интрафузальных мышц.
Вегетативные рефлексы – это ответная реакция внутренних органов. Эти рефлексы осуществляются за счет аксонов вегетативных нейронов, расположенных в боковых рогах спинного мозга и составляющих периферические нервы симпатического и парасимпатического отделов АНС. Эти рефлексы участвуют в регуляции артериального давления, деятельности сердца, секреции и моторной функции пищеварительного тракта и функции мочеполовой системы.
Проводниковая функция спинного мозга: восходящие, идущие к различным отделам голвного мозга, и нисходящие – от головного мозга к спинному. К восходящим путям относится: 1) пучок Бурдаха (от рецепторов верхней части тела и в/конечностей) и Голя (от рецепторов нижней части тела и н/конечностей). Это пути тактильной чувствительности (прикосновение и давление). Импульсы по этим путям достигают таламуса и отсюда в заднюю центральную извилину (кора больших полушарий); 2) путь болевой и температурной чувствительности, или дорсальный спиноталамический путь – от ноцицепторов и терморецепторов к канатиковым интернейронам, отсюда на противоположную сторону и до таламуса; 3) дорсальный (пучок Флексига) и вентральный спиномозжечковый пути передают информацию от сухожилий, кожи и висцерорецепторов до мозжечка. Эти пути участвуют в поддержании тонуса мышц при движении и сохранении позы тела в прострастве. К нисходящим путям относится: 1) пирамидный, или кортикоспинальный (латеральный и передний). Эти пути обеспечивают связь нейронов двигательной зоны КБП с альфа мотонейронами спинного мозгаи отвечает за произвольные движения; 2) руброспинальный (красноядерно-спиномозжечковый) тракт (Манакова) относится к экстрапирамидной системе, связывает нейроны красного ядра среднего мозга с мозжечком, продолговатым мозгом и спинным – управляет тонусом мышц и непроизвольной координацией движений; 3) вестибулоспинальный также относится к экстрапирамидной системе, отвечает за связь между ядром Дейтерса варолиевого моста, мозжечком и альфа мотонейронами передних рогов спинного мозга. Регулирует тонус мышц, координацию движений, равновесие и ориентацию в пространстве; 4) ретикулоспинальный путь – также относится к экстрапирамидной системе. В зависмости от того в каких нейронах спинного мозга заканчивается этот путь различают следующие функции: а) торможение спиномозговых рефлексов – при этом путь заканчивается на клетках Реншоу, их возбуждение приводит к гиперполяризации альфа мотонейронов, что приводит к торможению; б) облегчение спиномозговых рефлексов – при этом путь заканчивается на тормозных интернейронах, их возбуждение приводит к торможению клеток Реншоу, поэтому снимается тормозной эффект этих клетое на альфа мотонейроны, происходит облегчение спиномозговых рефлексов; в) повышает тонус скелетных мышц – при этом путь заканчивается на гамма мотонейронах спинного мозга, их возбуждение приводит к сокращению интрафузальных мышц, происходит натяжение ядерной сумки, возбуждение рецепторов, усиливается поток импульсов по афферентному пути через задние корешки спинного мозга в альфа мотонейроны, при возбуждении которых увеличивается тонус мышц.
Функции продолговатого мозга. Классификация тонических рефлексов по Магнусу.
Продолговатый мозг. Выполняет две функции: 1) рефлекторную и 2) проводниковую. Рефлекторная функция осуществляется благодаря: а) ядрам 8 пар черепномозговых нервов (VIII-XII); б) автоматическим центрам (дыхательный, сосудодвигательный, чихания, кашля, морганья, рвоты, пилорического); в) участие нейронов продолговатого мозга в тонических рефлексах (статические рефлексы). Тонические рефлексы обеспечивают регуляцию мышечного тонуса. Афферентные импульсы, вызывающие эти рефлексы приходят по вестибулярной ветви VIII пары черпномозговых нервов, по спиномозговым, проводящим импульсы от рецепторов мышц лица, шеи, конечностей и туловища. Все тонические рефлексы Магнус разделил на две группы: 1) статические – при раздражении рецепторов этого рефлекса обуславливают определеное положение тела в пространстве; 2) статокинетические – сохраняют равновесие тела в пространстве при его перемещении. Статические рефлексы в свою очередь можно разделить на две группы: 1) познотонические – обеспечивают определенную позу тела в пространстве; 2) установочные, или выпрямительные – обеспечивают возвращение тела из неестественного положения в нормальное (из горизонтального положения в положение стоя). Центр статических познотонических рефлексов находится в продолговатом мозге, а центр статических выпрямительных и статокинетических – в среднем мозге. Афферентны пути этих рефлексов начинаются с рецепторов вестибулярного аппарата (предверия улитки и полукружных канальцев), проприорецепторов мышц шеи и тактильных рецепторов кожи туловища сбоку. Значение проприорецепторов мышц шеи: при запрокидывании головы назад повышается тонус мышц разгибателей верхних конечностей и понижается тонус мышц разгибателей нижних конечностей; при нагибании головы к грудной клетке – понижается тонус мышц разгибателей верхних конечностей и повышается тонус мышц разгибателй нижних конечностей; при повороте головы влево – повышается тонус мышц разгибателей верхней кончности слева и повышается тонус мышц сгибателей справа; при повороте головы вправо – повышается тонус мышц разгибателей верхней кончности справа и повышается тонус мышц сгибателей слева. Выпрямительные рефлексы осуществляются в две фазы: 1) подъем головы – за счет раздражения рецепторов вестибулярного аппарата (предверия улитки) и кожной поверхности туловища; 2) выпрямление туловища – при раздражении проприорецепторов мышц шеи и кожной поверхности туловища. Статокинетические рефлексы осуществляются при вращательной пробе – при этом возникают вестибуломоторные, вестибуловегетативные и вестибулосенсорные рефлексы. Проводниковая функция продолговатого мозга – через него проходят все восходящие и нисходящие пути спинного мозга
Функции мозжечка. Мозжечок выполняет функцию координации и регуляции произвольных и непроизвольных движений. Эти функции обеспечиваются множественными связями мозжечка со спинным мозгом и другими структурами головного мозга: 1) спиноцеребелярный путь – информирует мозжечок о тонусе скелетных мышц; 2) веточка от кортикоспинального пути несет информацию о необходимом тонусе скелетных мышц; 3) церебелоспинальный – этот путь корегирует необходимый тонус скелетных мышц в конкретной ситуации (путем сравнения сигналов от спиноцребелярного пути и кортикоспинального). 1-3 пути обеспечивают координацию тела в пространстве при осознанных движений; 4) двухсторонняя связь мозжечка с продолговатым и средним мозгом – этот путь обеспечивает координацию неосознанных движений; 5) двухсторонняя связ с гипоталамусом и таламусом – обеспечивает координацию сенсорных и вегетативных функций.
При экспериментальном одностороннем поражении мозжечка наблюдаются следующие симптомы: 1) манежные движения – животное движется по кругу; 2) штопорообразные движения – если лягушку опустить в воду она совершает штопорообразные движения. Оба симптома связаны с тем, что при одностороннем поражении мозжечка происходит ярко выраженная дистония – на стороне поражения усиливается тонус мышц сгибателей и исчезает тонус мышц разгибателей, а на здоровой стороне наоброт: повышается тонус мышц разгибателей и исчезает тонус мышц сгибателей. При двухстороннем поражении отмечается полная обездвиженность, животное самостоятельно не может передвигаться.
В клинике отмечается частичное поражение мозжечка и различают следующие клинические симптомы: первые три симптома были описаны Лючиани и названы три а, так как все симптомы начинаются с буквы а: 1) астения – быстрая утомляемость мышц, так как при этом в двигательный акт вовлекаются «лишние» группы мышц за счет поражения 1-3 пути; 2) атония – отсутствие тонуса скелетных мышц, затем было установлено, что тонус мышц полностью не исчезает, а происходит нарушение тонуса (тонус одних мышц повышается, других понижается), поэтому првильнее этот симптом называть дистония; 3) астазия – утрачивается способность мышц к тетаническому сокращению – этот симптом проявляется в том, что при движении происходит качание различных частей тела; помимо вышеперечисленных симптомов наблюдаются 4) дизартрия – нарушение плавности речи; 5) дисметрия – нарушается соразмерность движения, как правило в сторону увеличения; 6) атаксия – нарушение координации движения, неуверенная походка; 7) эквилибристика – нарушение равновесия при ходьбе.
Функции ретикулярной формации мозга. Ретикулярная формация мозга – представленная диффузным скоплением клеток разного вида и величины, разделенных множеством разнонаправленных волокон. Этот отдел так был назван Дейтерсом в 1885 году за характерную цитоархитектонику. РФ располагается в центральной части мозгового ствола (между таламусом и спинным мозгом). Название ретикулярной формации, или сетевой, оно получило благодаря сетевых связей почти со всеми стуктурами ЦНС. Ретикулоспинальный путь относится к экстрапирамидной системе. Этот путь проводит импульсы от ретикулярной формации к различным нейронам спинного мозга. В зависмости от того в каких нейронах спинного мозга заканчивается этот путь различают следующие функции:
а) торможение спиномозговых рефлексов – при этом путь заканчивается на клетках Реншоу, их возбуждение приводит к гиперполяризации альфа мотонейронов, что приводит к торможению;
б) облегчение спиномозговых рефлексов – при этом путь заканчивается на тормозных интернейронах, их возбуждение приводит к торможению клеток Реншоу, поэтому снимается тормозной эффект этих клетое на альфа мотонейроны, происходит облегчение спиномозговых рефлексов;
в) повышает тонус скелетных мышц – при этом путь заканчивается на гамма мотонейронах спинного мозга, их возбуждение приводит к сокращению интрафузальных мышц, происходит натяжение ядерной сумки, возбуждение рецепторов, усиливается поток импульсов по афферентному пути через задние корешки спинного мозга в альфа мотонейроны, при возбуждении которых увеличивается тонус мышц.
Восходящие пути от РФ: 1) ретикулокортикальные пути – от центра Мэгуна Моруции – повышает тонус нейронов КБП – бодрствующее состояние; 2) связь с гипоталамусом и лимбической системой (миндалевидный комплекс, гипокамп и поясная извилина) – этот путь обеспечивает целостную реакцию организма и регулирует вегетативные и гомеостатические функции организма.
Функции гипоталамуса. Гипоталамус: серый бугор, мамилярные тела и серое вещество, образующее дно и стенки 3 желудочка. Обширная связь гипоталамуса с КБП, мозжечком, РФ, парсимпатическими ядрами продолговатого мозга, симпатическими ядрами спинного мозга, таламусом, гипофизом обеспечивает многообразие его функций, обеспечивающих 32 парами ядер, которые подразделены на пять групп: 1) ядра преоптической области; 2) передняя группа ядер; 3) наружняя группа ядер; 4) средняя группа ядер; 5) задняя группа ядер. К функциям гипоталамуса относится:
1) это высший центр АНС: задняя группа ядер повышает тонус симпатического отдела, а передняя группа ядер – тонус парасимпатического отдела (см. раздел АНС). Эти ядра принимают участие в формировании вегетативного компонента эмоций;
2) высший центр эндокринной функции – через систему либеринов и статинов регулирует выработку тропных гормонов, которые в свою очередь регулируют выработку горрмонов-эффекторов через эндокринные железы (см. раздел физиологии эндокринной системы). Либерины и статины участвуют в формировании эндокринного компонента эмоции;
3) участие в терморегуляции через центр терморегуляции, состоящий из двух отделов – центра теплоотдачи и центра теплопродукции (см. функциональную систему, обеспечивающую постоянство температуры тела);
4) участвует в регуляции сна и бодрствования. Здесь обнаружен центр Гесса, при возбуждении которого происходит торможение бодрствующего центра Мэгуна Моруции, что приводит к разлитому торможению КБП – возникновению сна;
5) участвует в регуляции пищеварения и формировании пищевой мотивации (см. физиологию пищеварения): раздражение наружних ядер (ц. голода) вызывает ощущение голода, раздражение вентромедиальных ядер (ц. насыщени) вызывает отказ от пищи;
6) за счет осморецепторов регулирует осмотическое давление крови через изменение диуреза (см. физиологию почек);
7) за счет гормонов-эффекторов вазопресина (антидиуретического) и окситацина участвует в регуляции диуреза, АД, сокращения матки и мышц альвеол молочной железы (окситацин);
8) является составной частью ЦНС, участвующей в формировании эмоций;
9) участвует в адаптационном поведении через различные биологические мотивации. Гипоталамус является пейцмеккером всех биологических мотиваций.
Нервная регуляция физиологических функций. Рефлекс. Особенности соматической и вегетативной дуги. Регуляция, или управление – это такие воздействия на систему, при которых система переходит из одного уровня функционирования на другой – заранее предусмотренный. Все воздействия на систему можно разделить на два вида:1) задающие сигналы – это такие воздействия, при которых система переходит на заданный уровень функционирования; 2) возмущающие сигналы – это такие воздействия, при которых происходит отклонение от заданного уровня функционирования (фактический уровень функционирования), их количество зависит от обстановки, в которой находится человек. Возмущающие сигналы отклоняют работу эффектора от заданного уровня, поэтому фактический результат работы эффектора всегда отличается от заданного. Отклонение фактического результата от заданного называется рассогласованием: чем меньше рассогласование, тем оптимальнее процессы регуляции.
Нервная регуляция - при этом воздействия на рабочий орган осуществляются через нервную систему, за счет рефлекса. Рефлекс – это ответная реакция организма при обязательном участии ЦНС. Рефлекс в переводе с латинского языка означает «отражение». Впервые термин «отражение» был применен Р.Декартом (1595 – 1650) для характеристики реакций организма в ответ на раздражение органов чувств. Материальной субстанцией, или структурной основой любого рефлекса является рефлекторная дуга – совокупность морфологических структур, которая обеспечивает осуществление рефлекса. Различают два вида рефлекторной дуги:
- соматическая, обеспечивающая ответную реакцию скелетных мышц в ответ на раздражение;
- вегетативная, обеспечивающая ответную реакцию внутренних органов и сосудов в ответ на раздражение.
Отмечаются некоторые особенности вегетативной рефлекторной дуги по сравнению с соматической: а) отростки афферентных нейронов в соматической рефлекторной дуге заканчиваются на альфа-мотонейронах в передних рогах спинного мозга, а в вегетативной – на вегетативных нейронах в боковых рогах спинного мозга; б) эфферентный путь в соматической рефлекторной дуге однонейронный, а в вегетативной – двухнейронный (преганглионарное волокно и постганглионарное); в) рабочий орган в соматической рефлекторной дуге скелетные мышцы, а в вегетативной – внутренние органы и сосуды.
Звенья рефлекторной дуги, их роль. Любая рефлекторная дуга состоит из пяти основных звеньев, выключение каждого из которых приводит к исчезновению рефлекса (рис.17):
1) рецептор (1) – звено, которое воспринимает раздражение и преобразует энергию раздражения в нервный импульс. Таким образом, можно сказать, что рецептор выполняет функцию переводчика: переводит сигналы раздражения в сигналы, понятные для ЦНС (нервный импульс); в рецепторе осуществляется примитивный анализ сигналов, о чем свидетельствует наличие специфических рецепторов. Так, рецепторы зрительного анализатора из множества раздражителей различает только световой раздражитель – это и есть примитивный анализ;
2) афферентный нервный путь (2) – передает нервный импульс от рецептора в ЦНС;
3) нервный центр (3) – здесь происходит высший анализ полученных сигналов в результате чего изменяется работа необходимого эффектора. Следует отметить, что нервный центр может иметь несколько вставочных нейронов, где происходит задержка импульса для анализа. Чем сложнее рефлекс, тем больше вставочных нейронов содержится в нервном центре;
4) эфферентный нервный путь ( 4) – обеспечивает переход сигнала от ЦНС к рабочему органу (эффектору). Эфферентный путь вегетативного рефлекторного пути (рис. 17 А) прерывается в ганглии и состоит из преганглионарного (4а) - и постганглионарного (4б) волокна;
5)
эффектор
(рабочий орган - 5 и 5а)). Таким образом,
соматический и вегетативный рефлекторный
путь состоит из 5 звеньев: рецептора,
афферентного пути, вставочного нейрона,
эфферентного пути и эффектора (рабочего
органа) Из вышеизложенного следует, что
при выключении любого звена будет
отсутствовать результат (ответная
реакция), хотя причины отсутствия
результата будут разными, так как каждое
звено выполняет только ей присущую
ф ункцию.
ункцию.
Рис. 17. Вегетативная (А) и соматическая (Б) рефлекторная дуга
Гуморальная регуляция. Звенья гуморальной регуляции. Гуморальная регуляция – это воздействия на органы и системы через жидкости (Humor) организма при действии на специфические рецепторы гормонов или биологически активные вещества (БАВ). Гормоны и БАВ могут выделяться в общее русло крови, но их конечный эффект определяется наличием в соответствующем органе специфических рецепторов. Таким образом, гуморальная регуляция осуществляется по принципу сигнала «SOS», то есть гормоны или БАВ проходят через все ткани, а реагируют только те из них, которые имеют специфические рецепторы. Отмечается ряд особенностей гуморальной и нервной регуляции. При нервной регуляции импульсы от рецептора по афферентным путям поступают в ЦНС, откуда по эфферентным путям идут к эффектору. Нервная регуляция осуществляется на базе рефлексов. При гуморальной регуляции импульсы от рецептора по афферентным путям поступают в ЦНС. При этом происходит возбуждение гипоталамуса, в результате чего выделяются либерины или статины, которые по автономному гипоталамо-гипофизарному тракту поступают в переднюю долю гипофиза – аденогипофиз. Здесь либо усиливается выделение соответствующего тропного гормона (при действии либеринов), либо – уменьшается (при действии статинов). Тропные гормоны гипофиза через кровь влияют на соответствующие эндокринные железы, которые выделяют гормон-эффектор, влияющие через кровь на функцию эффектора. Таким образом, при гуморальной регуляции в процесс включаются несколько промежуточных эффекторов (гипоталамус, гипофиз, эндокринные железы), поэтому скорость гуморальной регуляции намного меньше скорости нервной регуляции.
Отличия нервной и гуморальной регуляции. Нервная регуляции функций отличается от гуморальной:
1) филогенетическое отличие – в процессе развития организма вначале возникает гуморальная регуляция, а затем на базе гуморальной возникает нервная регуляция;
2) пути, по которым осуществляются эти виды регуляций – при гуморальной регуляции – это сосуды, а при нервной - это нервы;
3) по скорости – при гуморальной регуляции максимальная скорость может быть 0,5 м/с, а при нервной – скорость может достигать 120 м/с. В связи с этим нервная регуляци – это срочный вид регуляции, а гуморальный – поддерживающий вид регуляции;
4) по источнику воздействия – при нервной регуляции – это нервный импульс, а при гуморальной – гормоны и БАВ;
5) по точности регуляции – нервная регуляция – это точный вид регуляции, так как импульс от ЦНС доходит до конкретного органа. Гуморальная регуляция не имеет точного адресата, так как гормоны или БАВ поступают в кровь и разносятся по всем органам и тканям. Этот вид регуляции осуществляется по механизму сигнала «SOS».
Функциональные системы организма (ФУС), его звенья. ФУС – это динамическая организация деятельность которой направлена на совершение какого-либо конечного полезного приспособительного результата (КППР). Динамическая организация, так как количество эффекторов, участвующих в той или иной ФУС постоянно меняется, а если КППР в пределах нормы, то ФУС не функционирует. Функциональных систем в организме много, каждая из которых отвечает за какой-то полезный результат. Например, ФУС, обеспечивающая постоянство газов крови, отвечает за оптимальное содержание кислорода и углекислого газа в крови. Любая ФУС по П.К. Анохину состоит из следующих компонентов (рис.18)
Рис. 18. Звенья функциональной системы организма
1) конечный полезный приспособительный результат (КППР -1) – это ведущее звено ФУС, так как от степени отклонения конечного результата зависит формирование ФУС. Любой КППР (артериальное давление, температура тела, газы крови и т. д.) находится в организме на каком-то оптимальном уровне. При отклонении КППР от оптимального уровня (в сторону увеличения или уменьшения), начинает формироваться ФУС, работа которого направлена на возвращение КППР к оптимальному уровню;
2) специфические рецепторы (2) – для каждого КППР имеются специфические рецепторы, которые начинают возбуждаться при отклонении КППР от оптимального уровня;
3) афферентный путь (3а,, 3б) по которому идут сигналы в ЦНС, свидетельствующие об отклонении КППР от оптимального уровня. Различают афферентный нервный путь (3а) от специфических рецепторов к ЦНС) и гуморальный афферентный путь (3б) - непосредственное изменение КППР воздействует на ЦНС током крови;
4) ЦНС (а,б,в), который включает три уровня: а) специфический центр, который возбуждается при изменении какого-либо показателя; б) гипоталамус, который является высшим центром всех вегетативных и эндокринных функций; в) кора больших полушарий, при возбуждении которого включается внешнее звено ФУС – целенаправленное поведение, направленное на конкретный результат;
5) эффекторы - рабочие органы (Э, Эс). Заметим, что в ФУС, в отличии от рефлекторной дуги, несколько эффекторов. Количество эффекторов в ФУС зависит от степени отклонения КППР от оптимального уровня и постоянно изменяется (в связи с этим ФУС – это динамическая организация, так как количество эффекторов в ней постоянно меняется). От специфического центра и гипоталамуса по эфферентным путям импульсы поступают на эффекторы, работа которых изменяет данный КППР. При вовлечении в процесс гипоталамуса параллельно с нервными импульсами включается гуморальное звено (Эс) воздействия на эффекторы через изменение функций эндокринных желез;
6) поведение (9)– это внешнее звено ФУС. Это звено включается в работу после того, как все эффекторы, входящие в данную ФУС, задействованы, но КППР не возвращается к оптимальному уровню. Таким образом, внешнее звено ФУС начинает функционировать после исчерпания внутренних резервов организма. Первый признак исчерпания внутренних резервов – это появление отрицательных эмоций. С этого момента начинает включаться в работу ФУС высший уровень ЦНС – кора больших полушарий, возникает система осознанных двигательных актов, которая проявляется в том или ином поведении. При этом поведение направлено на удовлетворение внутренней потребности организма. Поведение будет меняться один за другим пока КППР не восстановится до оптимальной величины.
Звенья центрального блока ФУС. Центральный блок любой ФУС (рис. 19) состоит из сследующих звеньев:
I.Афферентного синтеза (АС), где объединяются сигналы нескольких порядков: а) пусковой сигнал (1), который поступает от специфического рецептора и свидетельствует об отклонении КППР; б) обстановочные сигналы (2), которые несут информацию об обстановке в которой находится объект через соответствующие рецепторы; в) сигналы из блока памяти (П); г) сигналы из доминирующего очага возбуждения (Д). При объединении вышеизложенных сигналов АС отвечает на следующие вопросы: 1) что делать? При объединении пускового и обстановочных сигналов; 2) как делать? При объединении пускового, обстановочных сигналов и поступающих из блока памяти; 3) когда делать? При объединении пускового, обстановочных сигналов, поступающих из блока памяти и доминирующего очага возбуждения;
II. Принятия решения (ПР) – включить в работу необходимые эффекторы
III. Программы действия (ПД), то есть какие эффекторы и в каком режиме должны функционировать, чтобы вернуть КППР на оптимальный уровень;
IV. При реализации ПД формируется (4) четвертый блок – акцептор результатов действия (АРД) – этот блок П.К. Анохин назвал аппаратом будущего предвидения. В этом блоке отражены параметры будущего функционирования эффекторов, то есть какими должны быть параметры функционирования эффекторов. При изменении функции эффекторов сигналы об их фактическом результате по обратной связи (5) поступают в АРД. В этом блоке происходит сравнение заданного результата с фактическим, поступающим по обратной связи. Если фактический результат не совпадает с заданным импульсы от АРД идут в ПД и при этом происходит изменение уже функционирующих эффекторов.
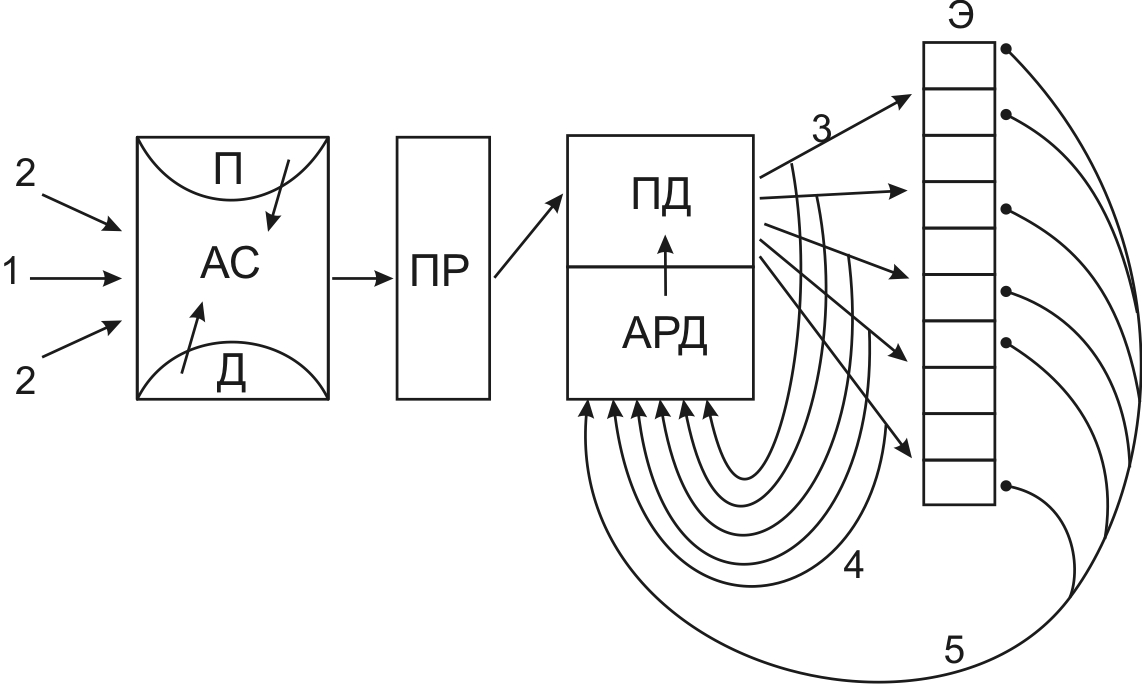
Рис. 19. Звенья центрального блока ФУС
Роль АС и АРД. Значение обратной связи в работе ФУС. Афферентного синтеза (АС) это первое звено центрального блока любой ФУС, где объединяются сигналы нескольких порядков: а) пусковой сигнал, который поступает от специфического рецептора и свидетельствует об отклонении КППР; б) обстановочные сигналы, которые несут информацию об обстановке в которой находится объект через соответствующие рецепторы; в) сигналы из блока памяти; г) сигналы из доминирующего очага возбуждения. При объединении вышеизложенных сигналов АС отвечает на следующие вопросы: 1) что делать? При объединении пускового и обстановочных сигналов; 2) как делать? При объединении пускового, обстановочных сигналов и поступающих из блока памяти; 3) когда делать? При объединении пускового, обстановочных сигналов, поступающих из блока памяти и доминирующего очага возбуждения.
Акцептор результатов действия (АРД) – это звено центральной части ФУС, который формируется при реализации программы действия. Это звено П.К. Анохин назвал аппаратом будущего предвидения. В этом звене отражены параметры будущего функционирования эффекторов, то есть какими должны быть параметры функционирования эффекторов. При изменении функции эффекторов сигналы об их фактическом результате по обратной связи поступают в АРД. В этом блоке происходит сравнение заданного результата с фактическим, поступающим по обратной связи. Если фактический результат не совпадает с заданным импульсы от АРД идут в ПД и при этом происходит изменение уже функционирующих эффекторов. Значение обратной связи заключается в том, что сигналы по этой связи поступают в АРД и информируют о фактическом результате работы эффекторов (в АРД заложены параметры работы эффекторов, которые должны быть).