

2 курс / Нормальная физиология / Общие_свойства_возбудимых_тканей_
.pdfдить больше 200 импульсов в секунду, т. е. функциональная лабильность синапсов не превышает 200 Гц.
4.Высокая избирательность к химическим веществам обусловлена специфичностью хемоpецептоpов постсинаптической мембраны.
5.В отличие от нервного волокна, где частота проведения импульса соответствует частоте его возникновения, для синаптической передачи характерна трансформация ритма импульсов.
Такое изменение частоты обусловлено двумя причинами: 1) низкая функциональная лабильность синапса — при частоте импульсов на пресинаптической мембране выше 200 Гц часть импульсов просто не проводится; 2) возникновение импульса возбуждения в центральных синапсах является результатом пространственной и временной суммации всех возбуждающих и тормозных постсинаптических потенциалов.
6. Способность к формированию следовых процессов (трениров-
ка синапсов) — эта уникальная способность синапсов хранить след предыдущих возбуждений. Суть этого явления заключается в том,
что чем интенсивнее и чаще работает синапс, тем легче и быстрее осуществляется его работа.
В основе тренировки синапсов лежит механизм посттетанической потенциации, которая обусловлена тремя основными факторами: 1) накопление ионов кальция в пресинаптической мембране; 2) активация синтеза медиатора; 3) активация синтеза мембранных рецепторов.
7.Чувствительность к О2 — потому, что необходима АТФ для восстановления МПП, для восстановления градиента концентрации по всем ионам, включая ионы Са++.
8.Чувствительность к наркотическим и психотропным веществам, алкоголю, токсинам обусловлена тем, что ионные каналы и рецепторы — это белки, которые могут вступить во взаимодействие
сдругими белками.
Задания для самостоятельной работы
1.Объясните что такое медиатор
2.Нарисуйте схему рефлекторной дуги и обозначьте на ней места синаптических контактов.
71
3.Нарисуйте схему синапса, назовите его компоненты, объясните:
a)механизм выделения медиатора;
b)роль ионов кальция в проведении нервного импульса;
c)физиологическую роль мембранных рецепторов постсинаптической мембраны;
d)разницу между метаботропными и ионотропными рецептора-
ми;
e)механизм изменения потенциала постсинаптической мембра-
ны.
4.Перечислите известные вам медиаторы.
5.Опишите механизмы действия синаптоактивных веществ.
6. Нарисуйте два графика: изменение потенциала постсинаптической мембраны при выделении ацетилхолина и глицина.
7. Дайте сравнительную характеристику проведению нервного импульса по нервному волокну и в химическом синапсе.
8. Какое отношение к бессоннице имеет посттетаническая потенциация синапсов?
Распространение возбуждения в нервных центрах
Теперь, когда нам известны механизмы проведения импульса по нервному волокну и через синапс, рассмотрим особенности прове-
дения информации через нервные центры, потому что именно здесь происходит самое существенное видоизменение нервного импульса. Вспомним Павловский принцип рефлекторной деятельности: рефлекторная реакция осуществляется на основании анализа и синтез информации. Это происходит именно в нервных центрах.
Нервные центры представляют собой совокупность нейронов, расположенных в одной или нескольких структурах ЦНС и регулирующих определенную функцию организма. Локализация и функция некоторых центров представлена в табл. 3.
Отметим принципиальные положения функционирования нервных центров: нервные центры собирают информацию от большого числа рецептивных полей — афферентных входов в нервном центре всегда намного больше, чем эфферентных выходов. В нервном центре формируется ответная реакция в виде импульса возбуждения и передается эффекторному органу.
72
Основные факторы, определяющие распространение возбуждения в нервном центре:
1.Проведение осуществляется синаптически, со всеми особенностями такого проведения.
2.Нейроны в нервном центре разные: и тормозные, и возбуждающие — в зависимости от того, какой медиатор синтезируется в нейроне.
3.Нейронов в нервном центре много. Афферентные нейроны получают сенсорную информацию и передают ее в центр, эти нейроны всегда возбуждающие. В нервном центре очень много вставочных нейронов, именно эти нейроны и модулируют проведение импульса по нервному центру. Двигательные нейроны обеспечивают передачу информации эффектору, они, как правило, возбуждающие, за исключением некоторых нейронов вегетативной нервной системы.
4.Обработка информации в нервном центре осуществляется на основе пространственной и временной суммации возбуждающих и тормозных импульсов.
Рассмотрим подробнее влияние всех факторов на изменение распространения нервного импульса в нервных центрах.
Первый фактор, определяющий распространение возбуждения
внервном центре: передача нервного импульса осуществляется синаптически. Все особенности синаптической передачи импульса мы обсудили в предыдущем разделе. Они в свою очередь определяют и свойства нервных центров: одностороннее проведение импульса, низкую лабильность, чувствительность к недостатку кислорода, токсинам.
Со свойствами синапсов связана важная особенность работы нервных центров – их пластичность. Пластичность — это способность к изменению функциональных свойств нервного центра под влиянием длительных внешних воздействий. Если по определенной цепочке нейронов импульс возбуждения передается часто и подолгу, то в результате импульс проводится именно по этой цепочке и быстрее, и легче. Можно считать, что сформировался новый путь распространения импульса с облегченным проведением.
В основе пластичности лежат два основных механизма. Первый, функциональный, мы уже обсудили — это результат «тренировки синапсов», в основе которого лежит посттетаническая потенциа-
73
ция синапса, которая может быть связана с накоплением в клетке ионов кальция, увеличением скорости синтеза медиатора и мембранных рецепторов. Второй, морфологический механизм заключается
втом, что на дендритах нейронов могут образовываться дендритные шипики как адаптивная реакция в ответ на интенсивную работу. В результате появления дендритных шипиков образуются новые, аксо-дендритных синапсы, а это уже структурная основа для нового пути связи между нейронами. Именно пластичность нервных центров, обусловленная свойствами синапсов, лежит в основе возможности образования новых связей между центрами различных рефлексов, формирования условных рефлексов, возможности обучаться, запоминать и хранить информацию. Пластичность центров чрезвычайно важна и в процессах компенсаторной перестройки (при повреждении центров).
Второй фактор, определяющий распространение возбуждения
внервном центре, заключается в том, что нейроны разные: в ЦНС есть возбуждающие и тормозные нейроны. Механизмы торможения играют большую роль в работе ЦНС, поэтому остановимся на них подробнее.
Механизмы торможения в ЦНС
Изучив принципы рефлекторной регуляции функций, процессы возникновения и проведения возбуждения, вы поняли, что на все эти процессы можно воздействовать: возможно блокировать проведение импульса новокаином; увеличить или уменьшить возбудимость ткани, изменив ионный состав окружающей среды; воздействовать синаптоактивными веществами. Однако есть механизм, с помощью которого сам организм воздействует на процессы возбуждения и проведения импульса — торможение.
Суть этого великолепного достижения эволюции необыкновенно проста: первый вариант — снижение возбудимости клетки вплоть до полной ее невозбудимости; второй вариант — изменение частоты идущих к клетке возбуждающих импульсов вплоть до прекращения проведения.
74
Механизмы торможения — это преподнесенный нам в руки природой еще один путь воздействия на процессы возбуждения и проведения нервного импульса: можно воздействовать на процессы торможения, но для этого их нужно хорошо понять.
СЛОВАРЬ ТЕМЫ:
—Тормозные нейроны.
—Тормозные медиаторы.
—ВПСП, ТПСП.
—Пресинаптическое торможение, постсинаптическое торможение.
—Первичное торможение, вторичное торможение.
—Торможение Введенского (пессимальное).
NaCl
H2SO4 |
H2SO4 |
Рис. 26. Схема опыта И.М.Сеченова для демонстрации центрального торможения до и после аппликации на зрительные бугры NaCl
75
Под торможением понимают самостоятельный нервный активный процесс, вызываемый возбуждением, который проявляется в угнетении или полном выключении другого возбуждения
Впервые возможность влияния на рефлекторные процессы без воздействия на структуры рефлекторной дуги была обнаружена И. М. Сеченовым. В 1863 г. И. М. Сеченовым было открыто централь-
ное торможение (Сеченовское).
И. М. Сеченову удалось установить, что сверхпороговое возбуждение центров среднего мозга приводит к торможению периферических, спинальных рефлексов.
Различаютпервичноеивторичноеторможение.Первичноетор-
можение обусловлено нервными импульсами, возникающими в особых тормозных нейронах.
Вцентральнойнервнойсистеметормозные нейроны есть в спин-
ном мозге, в головном мозге (меньшее количество) и в коре головного мозга (большинство).
Вспинном мозге два вида тормозных нейронов (это тормозные вставочные нейроны):
— клетки Реншоу: не обладают фоновой активностью и в покое не генерируют нервных импульсов. Они возбуждаются под действием: импульсов от афферентных нейронов, от эфферентных нейронов (альфа-мотонейронов спинного мозга), импульсов от вышележащих отделов головного мозга;
— клетки Уилсона: обладают постоянной фоновой активностью, даже без раздражения (в покое) они генерируют нервные импульсы — постоянно тормозят активность альфа-мотонейронов спинного мозга.
Вголовном мозге существуют отдельные клетки Реншоу и клетки Пуркинье, грушевидные нейроны мозжечка: они оказывают торможение внутри мозжечка, ядер среднего и продолговатого мозга, тем самым обеспечивается правильное распределение мышечного тонуса.
Вкоре головного мозга 4 вида тормозных клеток:
— большие корзинчатые нейроны: 3, 4, 5 слои коры головного мозга, их аксоны сильно ветвятся и образуют сплетения на площади около 500 мкм. Они тормозят активность нейронов 3, 4, 5 слоев;
76

—малые корзинчатые клетки нейроны: 2, 3 слои коры, их ак-
соны ветвятся на меньшей площади, около 50 мкм, и тормозят 2 и 3 слои;
—нейроны с кистеобразным аксоном: 1 слой коры, образует
аксон, на конце которого разветвление в виде кисти; тормозит клетки 1-го слоя;
— канделяброобразные нейроны: на границе между 2 и 3 слоя-
ми, вниз от них идет аксон и дает несколько ответвлений вверх, тормозят активность всех слоев.
Первичное торможение осуществляется за счет выделения тормозного медиатора на окончаниях нервных клеток (ГАМК — гаммааминомаслянная кислота, глицин, таурин, серотонин и другие). Эти вещества вызывают гиперполяризацию постсинаптической мембраны и, как следствие, тормозной постсинаптический потенциал.
Вспомним результат выделения медиатора пресинаптической мембраной: на постсинаптической мембране происходит деполяризация и возникает локальный ответ. Тормозные нейроны синтезиру- ютдругиемедиаторы—тормозные.Эти медиаторы, взаимодействуя со своими рецепторами, открывают другие каналы, например калиевые или хлорные. Что же происходит в этом случае с постсинаптической мембраной?
возбуждающий синапс |
тормозной синапс |
Ацетилхолин |
ГАМК, глицин |
открываются Na-каналы — депо- |
открываются К-каналы — гипер- |
ляризация |
поляризация |
Тормозной медиатор, как и возбуждающий, активирует рецептор управляемые каналы, однако не натриевые, а калиевые. Эта активация вызывает увеличение тока ионов калия. Если вспомнить о том, что ионы калия движутся по градиенту концентрации из клетки, то
77
становится ясно, что при активации калиевых каналов происходит гиперполяризация мембраны и снижение возбудимости нейрона.
Изменение потенциала мембраны при гиперполяризации называ-
ется тормозной постсинаптический потенциал (ТПСП). Во мно-
гих тормозных синапсах аналогичный эффект имеет активация каналов для хлора, увеличивающая его транспорт внутрь клетки. Результатом ТПСП является удаление МПП от критического уровня деполяризации, возбуждение становится либо вовсе невозможным, либо для него требуется значительно большая сила раздражителя.
Нервные импульсы, возникающие при возбуждении тормозных нейронов, вызывают гиперполяризацию постсинаптической мембраны и тормозной постсинаптический потенциал (ТПСП), это наиболее распространенный тип торможения в нервной системе, кото-
рый называется постсинаптическим торможением. Постсинапти-
ческое торможение является первичным торможением — самостоятельный процесс, вызванный возбуждением тормозных нейронов.
Центральные синапсы отличаются тем, что медиаторы вызывают появление на постсинаптической мембране только локального ответа, (ВПСП или ТПСП). ВПСП обладает всеми свойствами ЛО — зависит от силы раздражителя (количества выделившегося медиатора), суммируется и не передается по аксону.
Амплитуда одиночного ВПСП довольно мала — несколько мВ, поэтому для уменьшения заряда мембраны до критического уровня необходима либо одновременная активация нескольких возбуждающих синапсов — пространственная суммация, либо повышение частоты поступающих импульсов — временная суммация.
Если одновременно с ВПСП на мембране нейрона формируется и ТПСП, то эти изменения мембранного потенциала суммируются, следовательно, если величины ВПСП и ТПСП примерно одинаковы, то изменения мембранного потенциала нейрона, на котором суммируются воздействия, не произойдет (рис. 27, 28).
На рис. 27 представлена схема регистрации мембранного потенциала возбуждающего и тормозного нейронов, которые одновременно воздействуют на один нейрон. И возбуждающий, и тормозной нейрон возбуждены — регистрируется ПД. Следовательно, одновременно выделяются и возбуждающий, и тормозной медиаторы. На
78
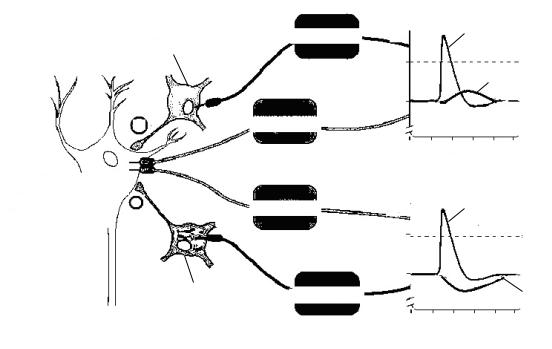
Суммация ВПСП и ТПСП МП не изменяется
Суммация
ВПСП и ТПСП усилитель МП не изменяется
усилитель
усилитель
Тормозной нейрон усилитель
+10 |
|
|
ПД возбуждающего |
|||
|
|
|
нейрона |
|
|
|
0 |
|
|
|
ВПСП |
|
|
mV |
|
|
|
|
||
|
|
|
|
|
|
|
−60 |
|
|
|
|
|
|
0 |
1 |
2 |
3 |
4 |
5 |
мс |
+10 |
|
|
ПД тормозного |
|||
|
|
|
нейрона |
|
|
|
0 |
|
|
|
|
|
|
mV |
|
|
|
|
|
|
−60 |
|
|
|
|
|
|
|
|
|
|
|
|
ТПСП |
0 |
1 |
2 |
3 |
4 |
5 |
мс |
Рис. 27. Суммация потенциалов на мембране нейрона
79

мембране тестируемого нейрона одновременно возникают ВПСП и ТПСП примерно одинаковой амплитуды, в результате суммации мембранный потенциал остается неизменным.
Временная суммация
Дендриты
Пространственная
суммация
Рис. 28. Виды суммации на нейроне
Существует еще один вариант первичного торможения в ЦНС: пресинаптическое торможение. Такое торможение возникает в том случае, когда нервный импульс или просто не достигает синаптического окончания аксона, или снижается его частота. Этот вид торможения осуществляется благодаря аксоаксональным синапсам, образованным терминалями аксонов тормозных интернейронов и аксонами возбуждающих нейронов (рис. 29, справа). В этих синапсах торможение определяется действием ГАМК, открывающей хлорные каналы на пресинаптических мембранах (мембрана возбуждающего нейрона). В результате на постсинаптической мембране тормозного синапса возникает явление стойкой деполяризации.
Деполяризация мембраны аксона ведет к инактивации натриевой проводимости. Возникает блокирование проведения нервного им-
80