

2 курс / Нормальная физиология / Методичка по физиологии
.pdf10.Морфо-функциональные особенности гладких мышц.
Гладкие мышцы обеспечивают механическую активность внутренних органов. Следовательно, их сокращения, по сравнению с сокращениями скелетных мышц:
-более медленные и длительные (нет необходимости в быстрых коротких движениях);
-менее координированные (нет необходимости в точных движениях);
-непроизвольные.
Существует два типа гладких мышц:
-унитарные;
-мультиунитарные.
Унитарные гладкие мышцы состоят из тесно связанных между собой клеток образующие гладкомышечные пласты. Клетки в пределах этих пластов соединены высокопроницаемыми межклеточными контактами – щелевыми контактами (нексусами), позволяющими ПД и местным потенциалам передаваться от одной клетки к другой. Такие пласты мышечных клеток в функциональном отношении ведут себя как синцитий и потому называются функциональным синцитием. Нервные волокна иннервируют лишь часть клеток пласта, а возникающий в этих клетках ПД передаются к остальным клеткам через щелевые контакты.
Особенности иннервации унитарных гладких мышц.
Вегетативные нервные волокна не образуют на гладкомышечных клетках типичных синапсов. В случае унитарных гладких мышц нервное окончание широко разветвляется над мышечным пластом, причем между нервными волокнами и мышечными клетками остается довольно большое расстояние. В ветвлениях нервного окончания имеются варикозные расширения, в которых скапливаются пузырьки с медиатором; при возбуждении нервного окончания медиатор высвобождается из всех варикозных расширений, изливаясь на большую поверхность гладкомышечного пласта. При этом до многих клеток (особенно в глубине гладкомышечных пластов) медиатор не доходит и ПД передаются к ним через щелевые контакты. Такие мышцы имеются в подавляющем большинстве внутренних органов, характеризующихся медленными генерализованными сокращениями (например, в органах ЖКТ), поэтому унитарные гладкие мышцы называются также висцеральными.
Мультиунитарные гладкие мышцы состоят из отдельных мышечных клеток, разделенных прослойками межклеточного вещества. Эти клетки сокращаются независимо друг от друга, и каждая из них имеет отдельную иннервацию, причем с каждой клеткой контактирует по меньшей мере одно варикозное расширение. Из таких мышц состоят органы, сокращения которых должны быть относительно быстрыми и точными, например, мышцы зрачка.
91
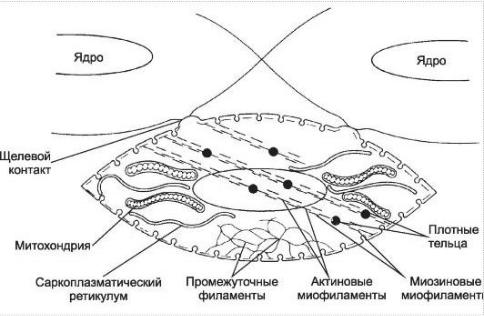
Рис. 24. Сократительный аппарат гладкомышечной клетки.
Морфологические особенности.
1.Образованы гладкомышечными клетками веретенообразной
формы.
2.Хаотично расположены и окружены соединительной тканью.
3.Контактируют друг с другом при помощи нексусов.
4.Сократительный аппарат представлен миофибриллами, однако:
-вместо упорядоченных миофибрилл со строгим чередованием нитей актина и миозина имеются пучки актиновых нитей, в центре которых располагается миозин;
-эти пучки не располагаются параллельно оси волокна, но прекрепляются к его мембране в области так называемы плотных телец; при сокращении актиновые пучки стягивают мембраны в области плотных телец, тем самым как бы сморщивая клетку;
-саркоплазматический ретикулум развит слабо, так как в сокращении участвует внеклеточный кальций.
Физиологические свойства.
Гладкие мышцы, как и скелетные, обладают возбудимостью, проводимостью и сократимостью. Однако эти свойства у глаких и скелетных мышц существенно отличаются.
Потенциал покоя в гладких мышцах меньше, чем в скелетных. В клетках не обладающих автоматией он стабилен и равен -60 -70 мВ. В клетках, обладающих автоматией, он неустойчивый с колебаниями от -30 до -70 мВ. Это связано с более высокой проницаемостью мембраны для ионов натрия.
92
Потенциал действия имеет длительный латентный период. Его амплитуда ниже, чем в скелетных мышцах. Бывает двух типов: пикоподобная форма и форма «плато». Быстрых натриевых каналов, отвечающих за ПД в нейронах и скелетных мышцах, в гладкомышечных клетках практически нет, и в любом случае при столь низком потенциале покоя они были бы полностью инактивированы. ПД в этих клетках формируется за счет входящего тока ионов Са2+ по так называемым
медленным кальциевым каналам. Эти каналы во много похожи на быстрые натриевые каналы:
-у них имеются наружные активационные ворота и внутренние инактивационные;
-активационные ворота в покое закрыты, при деполяризации открываются;
-инактивационные ворота в покое открыты, при деполяризации
закрываются.
Отличия медленных кадьциевых каналов от быстрых натриевых следующие:
-они пропускают преимущественно кальций;
-их ворота срабатывают значительно медленнее, то есть медленнее открываются и медленнее закрываются.
Всвязи с медленной работой ворот кальциевых каналов ПД гладкомышечных клеток отличается:
-меньшей крутизной;
-меньшей амплитудой;
-большей длительностью (десятки миллисекунд).
Автоматизм. Некоторые гладкомышечные клетки обладают свойством автоматизма – т.е. способность самопроизвольно без внешних раздражителей генерировать ПД. У этих клеток нет потенциала покоя, но имеются так называемые медленные волны, представляющие собой медленную самопроизвольную деполяризацию. Когда эта деполяризация достигает КУД, возникает один или несколько ПД.
Клетки (или группы клеток) обладающие способностью к автоматизму называются водителями ритма или пейсмекерами.
Генерация ПД. В скелетных мышцах ПД возникает только под действием импульса, поступающего по аксону мотонейрона. В гладких мышцах ПД может возникнуть:
-самопроизвольно (в клетках обладающих автоматизмом);
-в результате поступления ПД от соседних клеток (через нексусы);
-под действием нервных и гуморальных влияний;
-в некоторых клетках – при их растяжении.
Нервные и гуморальные влияния, а также растяжение гладкомышечных клеток часто приводят к генерации ПД не сами по себе, а на фоне медленных волн; эти влияния приводят к деполяризации клетки
93
и тогда Екр (или КУД) в результате очередной медленной волны достигается легче.
Проводимость. В унитарных гладких мышцах ПД проводятся не только вдоль всей мембраны клетки, но и от одной клетки к другой через щелевые контакты, охватывая большие мышечные пласты. Распространяется ПД лишь на определенное расстояние, которое зависит от силы раздражителя. Скорость распространения возбуждения составляет от 2 до 15 см/сек.
Сократимость. Общие принципы сокращения едины для всех типов мышц. В то же время сокращение гладких мышц обладает особенностями на всех его этапах:
-источниками кальция;
-механизмом пускового действия кальция;
-энергетикой сокращения;
-механизмом расслабления.
Источники Са2+. В гладкой мышце СПР выражен слабо, и поэтому главный источник Са2+ - вход его из внеклеточной среды по медленным кальциевым каналам сарколеммы. Поступление из СПР – дополнительный, но не основной источник.
Механизм действия Са2+. В гладкой мышце, как и в скелетной, Са2+ запускает взаимодействие актина с миозином через регуляторные белки. Однако, и сами эти белки и их действия совершенно иные.
1.Са2+ связывается с белком кальмодулином.
2.Комплекс кальций-кальмодулин активирует фермент киназу легких цепей миозина.
3.Этот фермент фосфорилирует миозин.
4.Только после этого поперечный мостик миозина приобретает способность связываться с активными центрами актина.
Энергетика сокращения. Главное отличие гладких мышц
заключается в низком сродстве миозина к АТФ, следовательно миозиновые мостики в течение долгого времени остаются соединенные с актином.
Механизм расслабления. Са2+ удаляется в основном во внеклеточную среду посредством кальциевого насоса сарколеммы и лишь в небольшой степени в СПР посредством кальциевого насоса ретикулума. После удаления Са2+ миозин остается фосфорилированным, а следовательно сохраняет способность взаимодействовать с актином. Дефосфорилирование миозина осуществляется фосфатазой легких цепей миозина. Лишь после этого прекращается взаимодействие актина с миозином, и мышца расслабляется.
Основные отличия между механизмом запуска взаимодействия актина с миозином поперечно-полосатой и гладкой мышцах.
1. И в поперечно-полосатой и в гладкой мышце в состоянии покоя актин
94
с миозином не взаимодействуют. Однако:
-в поперечно-полосатой мышце это обусловлено тем, что активные центры актина закрыты тропомиозином;
-в гладкой мышце тропомиозина нет, активные центры открыты, но миозин не обладает сродством к актину.
2.В обеих мышцах взаимодействие актина и мозина запускается Са2+. Однако:
-в поперечно-полосатой мышце Са2+, соединяясь с тропонином вызывает открытие активных центров актина вследствие сдвига молекулы тропомиозина;
-в гладкой мышце Са2+ через ряд промежуточных этапов (образование кальций-кальмодулинового комплекса, активации киназы легких цепей миозина, фосфорилирования миозина) придает миозину сродство к актину.
3.В обеих мышцах для прекращения взаимодействия актина с миозином Са2+ должен быть удален из саркоплазмы. Однако:
-в поперечно-полосатой мышце удаление Са2+ приводит к мгновенному закрыванию активных центров актина тропомиозином и, как следствие, к мгновенному прекращению сокращения и началу расслабления;
-в гладкой мышце удаление Са2+ приводит лишь к прекращению дальнейшего фосфорилирования миозина, однако уже фосфорилированный миозин продолжает взаимодействовать с актином. Для прекращения этого взаимодействия и начала расслабления надо, чтобы миозин был дефосфорилирован фосфатазой легких цепей миозина.
4.В поперечно-полосатой мышце АТФ участвует только в образовании поперечных мостиков: на каждый цикл поперечного мостика расходуется 1 молекула АТФ. В гладкой мышце АТФ также участвует в образовании поперечных мостиков, но помимо этого необходима для фосфорилирования миозина и сообщения ему сродства к актину. Таким образом, в гладкой мышце АТФ присоединяется к двум разным участкам: к миозиновым головками при образовании поперечных мостиков и к легким цепям миозина при фосфорилировании киназы легких цепей.
Особенности сокращения.
1.Медленное сокращение. Основная причина – низкое сродство миозина к АТФ, а, следовательно, большое время цикла поперечных мостиков.
2.Медленное расслабление. Причина: необходимо время для
дефосфорилирования миозина фосфатазой после удаления из саркоплазмы Са2+.
95
3.«Феномен защелки». Этот феномен заключается в том, что после развития напряжения гладкая мышца способна долго оставаться сокращенной, причем энергозатраты на это сокращение резко падают (мышца как бы «защелкивается» в сокращенном состоянии). Этот механизм позволяет гладкой мышце поддерживать длительное тоническое сокращение в течение нескольких часов при использовании незначительного количества энергии.
4.Пластичность. При растяжении гладкой мышцы она длительное время может сохранять растянутую форму, что важно для внутренних органов, выполняющих депонирующую функцию (желудок, мочевой и желчный пузыри).
11.Моторные синапсы, строение, функциональные свойства, механизм передачи возбуждения. Фармакологическая коррекция работы мионеврального синапса.
Основным способом передачи возбуждения между нервными и мышечными клетками является химический, который реализуется с помощью синапсов. Синапс — это специализированное по структуре и функции место контакта мембран между двумя возбудимыми клетками, которое обеспечивает передачу импульсов от одной возбудимой клетки к другой с помощью молекул химических веществ, или медиаторов. Проведение возбуждения между нервом и мышцей осуществляется через нервно-мышечный синапс, который представляет собой место контакта окончания аксона нейрона с мембраной мышечного волокна. Функцией такого соединения является передача сигналов возбуждения от а- мотонейронов спинного мозга к скелетным мышечным волокнам, вызывающих сокращение мышцы.
Нервно-мышечный синапс имеет общие для всех синапсов структурные элементы:
- пресинаптическое окончание; - постсинаптическую мембрану; - синаптическую щель.
Пресинаптическое окончание образуется расширениями по ходу разветвления аксона, иннервирующего мышечное волокно. В нервномышечном синапсе пресинаптическое окончание имеет большую длину (около 1—2 мм). Часть мембраны пресинаптического окончания, ограничивающая синаптическую щель, называется пресинаптической мембраной. Через нее осуществляется выделение (экзоцитоз) медиатора в синаптическую щель.
96

Рис. 25. Схема нервно-мышечного синапса.
Главным ультраструктурным фрагментом пресинаптического окончания являются синаптические пузырьки (везикулы) диаметром около 40 нм. Они образуются в комплексе Гольджи, с помощью быстрого аксонного транспорта доставляются в пресинаптическое окончание и там заполняются медиатором и АТФ. В пресинаптическом окончании содержится несколько тысяч везикул, в каждой из которых имеется от 1 до 10 тыс. молекул химического вещества, участвующего в передаче влияния через синапс и в связи с этим названного медиатором (посредником). В везикулах терминали аксона, иннервирующего мышцу, содержится медиатор ацетилхолин. Ацетилхолин синтезируется непосредственно в нервном окончании из холина и ацетил-Ко-А при участии фермента ацетилтрансферазы. Количество ацетилхолина, высвобождаемое из одной везикулы, принято называть квантом медиатора. Без ресинтеза ацетилхолина в двигательном нервном окончании запаса его молекул в синаптических пузырьках достаточно для проведения примерно 10 000 импульсов. Везикулы пресинаптической терминали находятся в активной и депонированной формах. Активные везикулы непосредственно участвуют в передаче возбуждения через нервно-мышечное соединение, т.е. в выделении медиатора в синаптическую щель. Молекулы медиатора депонированной формы везикул постоянно пополняют пул активной формы медиатора по мере их использования в синаптической передаче. Число везикул является постоянной величиной пресинаптической терминали и обусловливает функциональную лабильность синапса.
В нервно-мышечном синапсе везикулы преимущественно расположены вблизи периодических утолщений пресинаптической
97
мембраны, называемых активными зонами. В неактивном синапсе везикулы с помощью белка синапсина связаны с белками цитоскелета, что обеспечивает их иммобилизацию и резервирование. Напротив скопления «синаптических» пузырьков (кластеров) вблизи пресинаптической мембраны постсинаптическая мембрана образует глубокие складки. Каждой из них соответствует активная зона пресинаптической мембраны - желобок на ее внутренней поверхности, вдоль обеих сторон которого располагаются в ряд синаптические пузырьки. Очевидно, активные зоны и ассоциированные с ними пузырьки следует рассматривать как аппарат, специализированный для экзоцитоза, т.е. для выброса содержимого этих пузырьков в синаптическую щель.
Важными структурами пресинаптического окончания являются митохондрии, осуществляющие энергетическое обеспечение процесса синаптической передачи, цистерны гладкой эндоплазматической сети, содержащие депонированный Са2+, а также микротрубочки и микрофиламенты, участвующие во внутриклеточном передвижении везикул.
Синаптическая щель в нервно-мышечном синапсе имеет ширину в среднем 50 нм. Она содержит межклеточную жидкость и мукополисахаридное плотное вещество в виде полосок, мостиков, которое обеспечивает связь между пре- и постсинаптической мембранами и может содержать ферменты. Фермент, расщепляющий ацетилхолин, называется ацетилхолинэстеразой. Молекулы этого фермента синтезируются в мышечной клетке и фиксируются к ее сарколемме со стороны синаптической щели, а также к плотному веществу синаптическй щели.
Постсинаптическая мембрана — утолщенная часть клеточной мембраны иннервируемой клетки, имеющая ионные каналы, содержащие белковые рецепторы, способные связать молекулы медиатора, т.е. эта часть мембраны иннервируемой клетки является хемочувствительной. Ее особенностью в нервно-мышечном синапсе является наличие множества мелких складок, которые образуют слепые карманы, открывающиеся в синаптическую щель. Благодаря им резко увеличиваются площадь постсинаптической мембраны и количество ее рецепторов, которое в одном синапсе достигает 10—20 млн. Постсинаптическую мембрану нервно-мышечного синапса называют также концевой пластинкой. В постсинаптической мембране нервно-мышечного соединения расположены ионотропные N-холинорецепторы (чувствительны к никотину).
Механизм передачи возбуждения через нервно-мышечный синапс.
Исходя из структурной организации нервно-мышечного синапса, передачу возбуждения с нерва на мышцу можно разделить на два физиологических механизма: пресинаптический и постсинаптический.
98
Пресинаптический механизм передачи возбуждения с нерва на мышцу является электро-химическим и его функция заключается в трансформации числа нервных импульсов в количество выделяющегося из нервной терминали медиатора. Проведение возбуждения через нервномышечный синапс связано с функцией активных зон пресинаптической мембраны, которые представляют собой комплекс рядами расположенных Са2+-каналов, синаптических везикул, специализированных белков экзоцитоза и элементов цитоскелета (актиновые филаменты и микротрубочки), объединенные вокруг так называемых плотных тел синапса. Последние представляют собой участки утолщения пресинаптической мембраны, в области которых происходит секреция медиатора.
После того как нервный импульс достигает пресинаптического окончания в фазу деполяризации потенциала действия, в мембране окончания открываются потенциалзависимые кальциевые ионные каналы, и ионы Са2+ пассивно по градиенту концентрации поступают внутрь нервного окончания из внеклеточной среды. В момент открытия кальциевых ионных каналов в активных зонах пресинаптической мембраны формируются микродомены. Микродомен представляет собой область высокой концентрации ионов Са2+ вблизи устья кальциевого ионного канала. В момент появления микродомена белки мембраны везикулы способны связывать везикулярную мембрану с активной зоной пресинаптического окончания. При этом образуется специфический белковый комплекс, состоящий из везикулы и элементов активной зоны пресинаптической мембраны, которые связаны между собой специализированными белками экзоцитоза. Этот комплекс называется секретосомой. Формирование секретосомы является основным процессом, обеспечивающим экзоцитоз ацетилхолина в нервном окончании.
Белки экзоцитоза локализованы как на мембране везикул (синапсин, синаптотагмин, синаптобревин), так и на пресинаптической мембране (синтаксин, синапсоассоциированный белок). Белок синаптотагмин является кальциевым рецептором. После связывания ионов Са2+ с синаптотагмином и последующего взаимодействия синаптобревина с синапсоассоциированным белком и синтаксином пресинаптической мембраны происходит связывание везикулы с активной зоной пресинаптической мембраны. Экзоцитоз медиатора обеспечивается за счет АТФазной активности специфического цитозольного белка. При высвобождении ацетилхолина в синаптическую щель мембрана везикулы полностью сливается с пресинаптической мембраной и медиатор выходит в синаптическую щель через так называемую пору.
Диффузия ацетилхолина через синаптическую щель нервномышечного синапса.
99
Ацетилхолин путем экзоцитоза поступает в синаптическую щель и затем диффундирует к постсинаптической мембране в течение 1 мс. Во время диффузии ацетилхолина через синаптическую щель примерно 1/3 от выделившегося количества медиатора разрушается ферментом ацетилхолинэстеразой на ацетат и холин, а 2/3 молекул ацетилхолина достигает постсинаптической мембраны и связывается с N- холинорецепторами. Активность ацетилхолинэстеразы столь высока, что период полураспада молекул ацетилхолина не превышает 1 мс. Инактивация ацетилхолина в синаптической щели исключает многократное связывание одних и тех же молекул медиатора с холинорецепторами постсинаптической мембраны, что предотвращает возможность непрерывного сокращения мышц или так называемой мышечной тетании.
Постсинаптический механизм – хемо-электрический.
Постсинаптический механизм передачи возбуждения через нервномышечный синапс представляет собой трансформацию количества выделившегося ацетилхолина в число потенциалов действия, которые генерирует сарколемма мышечного волокна. В постсинаптической мембране нервно-мышечного соединения расположены ионотропные N- холинорецепторы (чувствительны к никотину). Молекулы ацетилхолина кратковременно (не более 2 мс) взаимодействуют с N- холинорецепторами, в результате в постсинаптической мембране открываются ионные каналы для катионов. Входящий ток ионов натрия преобладает над выходящим калиевым током, в результате происходит деполяризация постсинаптической мембраны нервно-мышечного синапса,
которая называется потенциалом концевой пластинки.
Продолжительность потенциала очень мала — не более 1—2 мс, а его амплитуда варьирует в зависимости от количества высвобождающегося ацетилхолина. При этом между амплитудой потенциала концевой пластинки и количеством высвобождающегося ацетилхолина имеется прямая зависимость.
Потенциал концевой пластинки электротонически распространяется вдоль мышечного волокна. При этом, если потенциал концевой пластинки достигает по величине порога возбуждения мышечной мембраны (30— 40 мВ), то он вызывает в ней генерацию распространяющихся потенциалов действия. В результате возбуждение с нервного окончания передается на мышечную клетку. Если пресинаптического окончания достигает один потенциал действия, то в синаптическую щель нервно-мышечного синапса высвобождается в среднем 100 квантов ацетилхолина. В этом случае возникает потенциал концевой пластинки, величина которого достаточна для того, чтобы вызвать генерацию потенциалов действия в мышечной мембране, прилегающей к концевой пластинке. Даже при снижении амплитуды потенциала концевой пластинки на 75% он
100