

2.1.1. Медленная спонтанная диастолическая деполяризация мсдд
Свойством МСДД (фаза 4 на рис. 1а) обладают Р-клетки (от англ. pacemaker водитель ритма), располагающиеся главным образом и синоатриальном узле (синусовом узле, СА-узле) и АВ-соединении.
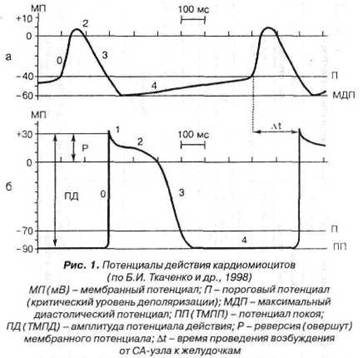
Фаза быстрой деполяризации Р-клеток (фаза 0) обусловлена вхождением Са+ и, частично, Na+ внутрь клеток через потенциалзависимые «медленные» мембранные каналы. Движение ионов против электрохимического градиента (активный транспорт) осуществляют ионные насосы, которые сопряжены с мембранными ферментами АТФ-азами. АТФ-азы ускоряют гидролиз АТФ, а выделяющаяся при этом энергия расходуется на перенос ионов. «Медленные» каналы открываются при величине трансмембранного потенциала около -40мВ. Фаза реполяризации «медленных» клеток (фазы 2 и 3) связана с инактивацией кальциевых каналов и увеличением выхода из клеток К+ (рис. 1а).
2.1.2. Трансмембранный потенциал покоя тмпп
Клетки рабочего миокарда и проводящей системы сердца в нормальных условиях не обладают свойством МСДД и в покое имеют стабильный ТМПП около -90мВ. Однако в условиях ишемии эти клетки могут «самовозбуждаться» и генерировать электрические импульсы.
ТМПП (рис. 1б) возникает вследствие того, что клеточная мембрана в состоянии покоя проницаема для К+ (в основном) и С1- (в меньшей степени). Во внутриклеточной среде концентрация К+ приблизительно в 30раз выше (140-150мМ), чем во внеклеточной среде (4-5мМ); а в противоположность этому Na+ в 20раз ниже, С1- в 13раз ниже, Са+ в 25раз ниже. Данное соотношение концентрации ионов поддерживается функционирующими ионными насосами. По направлению градиентов концентраций положительно заряженные ионы калия покидают клетку, а отрицательно заряженные ионы хлора поступают внутрь клетки. В результате этого наружная поверхность клеточной мембраны заряжается положительно, а внутренняя отрицательно (поляризация клеточной мембраны). Кроме того, в формировании диастолического потенциала кардиомиоцитов принимает участие и активный ток ионов, непосредственно создаваемый калиево-натриевым насосом. При работе этого насоса происходит неэквивалентный (электрогенный) обмен ионов: па каждые 2К+, введенных в клетку, выводится 3Na+. В результате возникает выходящий из клетки ток положительных зарядов - насосный ток, который увеличивает отрицательный внутриклеточный заряд.
2.1.3. Трансмембранный потенциал действия тмпд
Под действием возбуждающего влияния Р-клеток в «быстрых» клетках (кардиомиоцитах) начинают функционировать «быстрые» натриевые каналы (полностью открываются при величине потенциала около -60мВ), что приводит к лавинообразному входу Na+ в клетку. Входящий ток Na+ приводит к уменьшению трансмембранного потенциала (деполяризации). Когда потенциал мембраны понижается примерно до -40мВ, открываются потенциалзависимые «медленные» мембранные каналы, идентичные таковым в «медленных» клетках (Р-клетках). Через эти каналы осуществляется дополнительный «медленный» ток Са+ и Na+. «Быстрый» входящий ток натрия и «медленный» входящий ток кальция и натрия уменьшают трансмембраниый потенциал до нуля и перезаряжают мембрану (овершут) до +30мВ. При достижении этого значения трансмембранного потенциала «быстрые» натриевые каналы инактивирумтся, а «медленные» кальциевые и натриевые каналы продолжают функционировать, что обеспечивает в дальнейшем фазу «плато». В момент достижения положительного пика трансмембраниого потенциала открываются специфические мембранные каналы, через которые в клетку поступает С1-, обеспечивая фазу начальной быстрой реполяризации. При этом положительный потенциал мембраны уменьшается, что приводит к активации потенциалзависимых калиевых каналов. К+ начинает выходить из клетки. Динамическое равновесие между «медленными» входящими токами кальция и натрия и выходящим током калия обеспечивает временную стабилизацию трансмембранного потенциала (фазу «плато»).
По мере преобладания выходящего калиевого тока отрицательный потенциал мембраны постепенно возрастает, и в дальнейшем (при трансмембранном потенциале около -40мВ) «медленные» кальциевые и натриевые каналы закрываются, а преобладающим становится лишь выходящий калиевый ток, который и обеспечивает фазу конечной быстрой реполяризации кардиомиоцитов. Трансмембранпый потенциал при этом вновь достигает величины около -90мВ (рис. 1б).
Фазы ТМПД:
фаза 0 - фаза быстрой деполяризации клеток (10мс);
фаза 1 - фаза начальной быстрой реполяризации клеток;
фаза 2 - фаза «платою (200мс);
фаза 3 - фаза конечной быстрой реполяризации клеток;
фаза 4 - фаза ТМПП (диастола).
2.2. ХАРАКТЕРИСТИКА ОСНОВНЫХ ФУНКЦИЙ СЕРДЦА
2.2.1. ФУНКЦИЯ АВТОМАТИЗМА
Функция автоматизма — это способность кардиомиоцитов вырабатывать электрические импульсы при отсутствии внешних раздражителей.
Данной способностью обладают клетки-водители ритма (Р-клетки), основным свойством которых является МСДД (раздел 2.1.1.).
В сердце существует несколько центров автоматизма.
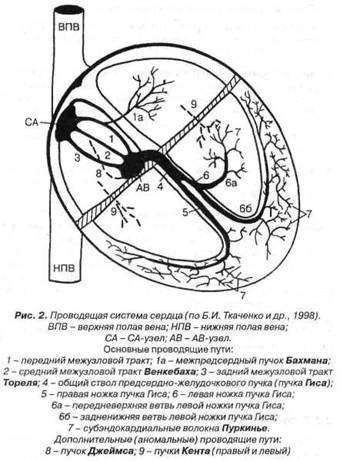
Центр автоматизма первого порядка - Р-клетки СА-узла, вырабатывающие импульсы у взрослых с частотой 60-80 мин, у детей до 140 мин и более.
Центры автоматизма второго порядка - Р-клетки верхней, срединно-боковой и нижней частей правого предсердия, стенок коронарного синуса и клапанов, верхней и нижнезадней стенок левого предсердия, АВ-соединения (в NH-зоне; кардиомиоциты АВ-узла функцией автоматизма не обладают). Эти клетки вырабатывают импульсы у взрослых с частотой 40-60 мин, у детей иногда с более высокой частотой.
Центры автоматизма третьего порядка - Р-клетки разветвлений системы пучка Гиса и волокон Пуркинье, вырабатывающие импульсы с частотой 20-40 мин.
Номотопным центром автоматизма (водителем ритма сердца, в обычных условиях) является СА-узел, расположенный в верхнезадней части правого предсердия между устьями полых вен (рис. 2). Степень автоматизма остальных (гетеротопных, латентных) центров автоматизма понижается по мере их удаления от СА-узла. В нормальных условиях автоматизм гетеротопных центров не проявляется, так как более частые синусовые импульсы, возбуждая все отделы сердца, разряжают зреющие в этих центрах импульсы.
Автоматизм латентных водителей ритма может проявиться:
при понижении синусового автоматизма;
при повышении автоматизма гетеротопного центра;
при блокаде на пути распространения синусового импульса.
2.2.2. ФУНКЦИЯ ВОЗБУДИМОСТИ
Функция возбудимости - это способность кардиомиоцитов возбуждаться под влиянием импульсов, исходящих от клеток-водителей ритма.
Эта функция подробно рассмотрена в разделе 2.1.3. и тесно связана с функцией проводимости (раздел 2.2.4.).
Нарушения функции возбудимости могут возникать:
при нарушении функции клеточных мембран;
при нарушении ионного баланса организма (калий, кальций, натрий, хлор и др.);
при неадекватном использовании лекарственных препаратов и др.
2.2.3. ФУНКЦИЯ РЕФРАКТЕРНОСТИ
Функция рефрактерности - это способность кардиомиоцитов временно не возбуждаться под влиянием импульсов, исходящих от клеток-водителей ритма. Выделяют:
абсолютный (эффективный) рефрактерный период (фазы 0, 1,2, начало фазы 3 ТМПД), когда кардиомиоциты не способны реагировать на импульс любой силы (раздел 2.1.3.);
относительный рефрактерный период (вторая половина фазы 3 ТМПД), когда сильные и сверхсильные импульсы способны преждевременно вызвать повторный ТМПД.
В заключительной стадии каждого цикла возбуждения сердца отмечают интервал времени, когда возникает период неоднородной рефрактерности кардиомиоцитов (у разных клеток различная скорость восстановления проводимости) - уязвимый период (период нестабильности миокарда).
Нарушения функции рефрактерности могут возникать при тех же обстоятельствах, что и нарушения функции возбудимости.