

2 курс / Гистология / ГИСТОФИЗИОЛОГИЯ_СОСУДИСТОЙ_ОБОЛОЧКИ_ГЛАЗА_ЧЕЛОВЕКА_В_ПРЕНАТАЛЬНОМ
.pdfСОВРЕМЕННОЕ СОСТОЯНИЕ ВОПРОСА ХОРИОИДАЛЬНОГО АНГИОГЕНЕЗА
рует пролиферацию Т-хелперов 2 (Th2) клонов, но не оказывает влияния на продукцию ими цитокинов. Антипролиферативное действие TNF-γ обусловлено экспрессией гена, ответственного за подавление выработки специфического пролиферативного фактора макрофагами. IL-2 индуцирует Т-клеточную пролиферацию, увеличивает число эффекторных клеток, потенцирует выделение других цитокинов. TNF-α, являющийся ключевой молекулой в воспалении, обладает антипролиферативным действием, которое обратимо только в течение 24 часов, в дальнейшем же происходит индукция апоптоза. Действие цитокинов необходимо рассматривать в связи с другими стимуляторами, прежде всего с гормонами, и с учетом типа клеток-мишеней и их тканевого микроокружения. Полипептиды, которые индуцируют дифференцировку и останавливают пролиферацию одних клеток, могут быть высокомитогенными для другого типа клеток [264, 341, 373].
Макрофаги принимают участие в регуляции пролиферативных процессов, т.к. в условиях культуры ткани они почти в 20 раз ускоряют рост клеток различных тканей. При этом макрофаги через моно- и лимфокины поддерживают прямые и обратные контакты с лимфоцитами. Активированные макрофаги, секретируя большой спектр цитокинов-монокинов (IL-1, IL-6, IL-8, IL-12, MIP-2, PDGF, TNF-α, G-CSF, GM-CSF), влияют на мигацию, пролиферацию и функцию моноцитов (предшественников макрофагов), нейтрофилов (IL-1, IL-8, MIP-2, PDGF, TNF-α, G-CSF, GM-CSF), Т- и В-лимфоцитов (IF-1, IL-6, IL-12, TNF-α) [151, 184, 279].
Решение вопроса о непосредственной секреции макрофагами ангиогенных факторов роста для эндотелия сосудов или посредничестве макрофагов для индукции секреции факторов роста другими клетками может помочь в решении задачи разработки методов ангиогенной консервативной терапии, предполагая индукцию ангиогенеза при введении факторов роста эндотелия или пула собственных моноцитов/макрофагов при ишемии в хориоидее [419]. Некоторые исследователи считают, что ряд заболеваний сосудистой системы глаза требует ингибирования ангиогенеза для нормализации зрительных функций [91, 101].
Под действием ангиогенных факторов тучные клетки мигрируют в участки ангиогенеза, где они также участвуют в его регуляции [202, 271]. Некоторые из этих цитокинов (сосудистый ростовой фактор эндотелиоцитов, интерлейкин-8 и др.) могут синтезироваться тучными клетками. Тучные клетки индуцируют морфологическую перестройку, вырабатывая интерлейкин-8, они привлекают в зону перестройки макрофаги [198, 220, 226, 331, 406].
Тучные клетки принимают участие в ангиогенезе опухолей с развитием неоваскуляризации [100, 303, 358, 360]. Гепарин, который могут
21

ГЛАВА I
синтезировать тучные клетки, вызывает пролиферацию эндотелиоцитов [174, 225, 307].
Кроме этого, тучные клетки способны сами синтезировать VEGF (vascular endothelial growth factor, фактор роста эндотелия сосудов). Ангиогенез при участии тучных клеток может происходить и без процессов дегрануляции [98, 271]. В тучных клетках располагаются гранулы с гистамином, которые способны стимулировать ангиогенез путем продукции VEGF [196, 292]. Кроме VEGF тучные клетки способны синтезировать bFGF (основной фактор роста фибробластов), который также влияет на ангиогенез [194, 306, 411].
Тучные клетки (Тк) кроме ангиогенеза обладают способностью вызывать пролиферацию фибробластов с формированием соединительной ткани [357, 359].
Особенности контактных взаимоотношений Тк с коллагеновыми волокнами предполагает стимуляцию ими пролиферативных процессов. Продуцируемые тучными клетками цитокины и факторы роста фибробластов, трансформирующий фактор роста способствуют не только васкуляризации формирующихся структур глаза, но и привлечению в зону перестройки тканей фибробластов и фиброкластов. С точки зрения регенерации повышенный интерес представляют клетки-предшественники тучных клеток, так как они являются прародителями клеток всех групп, участвующих в физиологической регенерации структур глаза [176, 248].
Специфика расположения тучных клеток позволяет предположить, что они имеют отношение к запуску перестройки структур и синтезу коллагеновых волокон. Знание механизма тучноклеточного цитокинового запуска перестройки структур глаза позволит по-новому взглянуть на современные концепции патогенеза глаукомы [81].
1.3. РАЗВИТИЕ ОРГАНА ЗРЕНИЯ
Филогенетический анализ описывает основные этапы становления органа зрения от отдельных светочувствительных клеток эктодермального происхождения (у кишечнополостных и плоских червей) до сложно устроенных парных глаз у млекопитающих. У позвоночных животных глаза развиваются из боковых выростов мозга, обеспечивающих формирование сетчатки. Средняя и наружная оболочки глазного яблока, стекловидное тело формируются из мезодермы (среднего зародышевого листка), хрусталик – из эктодермы [4, 65, 298].
Составные части глазного яблока, вспомогательный и защитные аппараты глаза развиваются из различных зачатков. Его светочувстви-
22

СОВРЕМЕННОЕ СОСТОЯНИЕ ВОПРОСА ХОРИОИДАЛЬНОГО АНГИОГЕНЕЗА
тельная часть – сетчатка – представляет собой часть стенки мозгового пузыря, выпячивающегося в середине 3-й недели внутриутробного развития (эмбрион имеет 7–8 сомитов) в виде двух пузырьков, полости которых сообщаются с полостью переднего мозгового пузыря [29, 66]. В начале 5-й недели дистальная часть глазного пузырька втягивается, что способствует уменьшению его полости и образованию двухслойной чаши. Внутренний слой чаши содержит клетки мозгового пузыря и представляет зачаток сетчатки, а наружные клетки относятся к эктодерме, дающей начало развитию хрусталика [61, 67, 316]. На 4–5-й неделях внутренний слой эктодермы в области глазной чаши утолщается и превращается в хрусталиковую пластинку, из которой развивается хрусталик. Из мезенхимы, окружающей глазной бокал, развиваются сосудистая
ибелочная оболочки [352, 353]. Мезенхима глазного яблока разделяется на внутреннюю пластинку – будущую сосудистую оболочку – и наружную плотную соединительнотканную пластинку – белочную оболочку. Передняя часть белочной оболочки формирует прозрачную роговицу, где наружный эпителиальный слой развивается из эктодермальных клеток [32, 297, 318].
На ранних этапах эмбрионального развития сосудистый бассейн стекловидного тела и кровеносные сосуды хориоидеи формируются также из мезенхимы, которая проникает в глазную чашу через сосудистую щель
ичерез щель между краями чаши и хрусталика [30, 53, 400]. Внутренняя оболочка (сетчатка) по форме напоминает двухстенный
бокал. Из тонкой наружной стенки бокала развивается пигментная часть (слой) сетчатки. Фоторецепторные светочувствительные клетки находятся в более толстом внутреннем слое бокала. В филогенезе у рыб дифференцировка зрительных клеток на палочковидные (палочки) и колбочковидные (колбочки) выражена слабо, у рептилий имеются одни колбочки,
умлекопитающих в сетчатке находятся преимущественно палочки;
уводных и ночных животных колбочки в сетчатке отсутствуют. В составе средней (сосудистой) оболочки уже у рыб начинает формироваться ресничное тело, усложняющееся в своем развитии у птиц и млекопитающих [66, 340, 393].
Умногих млекопитающих, кроме человека, имеется хориоидальное, или блестящее зеркальце (тапетум), которое образовано тапетацитами или коллагеновыми фибриллами и расположено на дне глаза, постепенно истончаясь в периферическом направлении [41, 42, 54].
Мышцы в радужке и в ресничном теле впервые появляются у амфибий. Наружная оболочка глазного яблока у низших позвоночных состоит преимущественно из хрящевой ткани (у рыб, частично у амфибий,
убольшинства ящерообразных и однопроходных). У млекопитающих она
23

ГЛАВА I
построена только из волокнистой (фиброзной) ткани. Передняя часть фиброзной оболочки (роговица) прозрачная. Хрусталик у рыб, амфибий округлый. Аккомодация достигается вследствие перемещения хрусталика
исокращения особой передвигающей хрусталик мышцы. У рептилий
иптиц хрусталик способен не только перемещаться, но и изменять свою кривизну. У млекопитающих хрусталик занимает постоянное место, аккомодация осуществляется вследствие изменения кривизны хрусталика. Стекловидное тело, имеющее вначале волокнистую структуру, постепенно становится прозрачным [67, 318, 372, 373].
Нельзя не отметить, что одновременно с усложнением строения глазного яблока развиваются вспомогательные органы глаза [342]. Первыми появляются шесть глазодвигательных мышц, преобразующихся из миотомов трех пар головных сомитов. Веки начинают формироваться у рыб в виде одной кольцевидной кожной складки. У наземных позвоночных животных образуются верхние и нижние веки, а у большинства из них у медиального угла глаза имеется также мигательная перепонка (третье веко). У обезьян и человека остатки этой перепонки сохраняются в виде полулунной складки конъюнктивы. У наземных позвоночных животных развивается слезная железа, формируется слезный аппарат [74, 75, 373, 432].
Глазное яблоко у человека также развивается из нескольких источников. Светочувствительная оболочка (СО, сетчатка) происходит из боковой стенки мозгового пузыря (будущий промежуточный мозг); главная линза глаза – хрусталик – непосредственно из эктодермы; сосудистая
ифиброзная оболочки – из мезенхимы. На ранней стадии развития зародыша (конец 1-го, начало 2-го месяца внутриутробной жизни) на боковых стенках первичного мозгового пузыря появляется небольшое парное выпячивание – глазные пузыри. Их концевые отделы расширяются, растут в сторону эктодермы, а соединяющие с мозгом ножки суживаются
ив дальнейшем превращаются в зрительные нервы. В процессе развития стенка глазного пузыря впячивается внутрь его, и пузырь превращается
вдвухслойный глазной бокал [77, 114,146, 160]. Наружная стенка бокала
вдальнейшем истончается и трансформируется в наружную пигментную часть (слой), а из внутренней стенки образуется сложно устроенная световоспринимающая (нервная) часть сетчатки (фотосенсорный слой). На стадии формирования глазного бокала и дифференцировки его стенок, на 2-м месяце внутриутробного развития, прилежащая к глазному бокалу спереди эктодерма вначале утолщается, а затем образуется хрусталиковая ямка, превращающаяся в хрусталиковый пузырек. Отделившись от эктодермы, пузырек погружается внутрь глазного бокала, теряет полость,
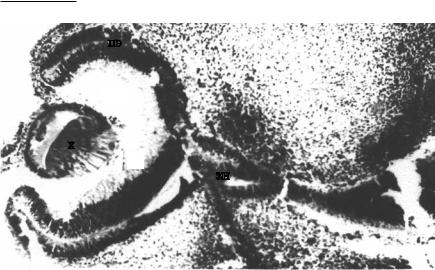
ииз него в дальнейшем формируется хрусталик (рис. 1) [167, 218, 374].
24
СОВРЕМЕННОЕ СОСТОЯНИЕ ВОПРОСА ХОРИОИДАЛЬНОГО АНГИОГЕНЕЗА
пэ
ст
х
с
зн
Рис. 1. Глазной бокал. Глаз эмбриона человека в возрасте 6 недель. Импрегнация серебром, ув. 100.
Условные обозначения: зн – зрительный нерв; пэ – пигментный эпителий; с – сетчатка; ст – стекловидное тело; х – хрусталик [80]. Фото из архива Г.В. Рева
Начиная со 2-го месяца эмбрионального развития через образовавшуюся на нижней стороне стенки глазного бокала щель начинают мигрировать мезенхимные клетки. За счёт этих клеток внутри бокала образуется кровеносная сосудистая сеть одновременно с формирующимся здесь стекловидным телом, расположенным вокруг растущего хрусталика. Материал распределяется таким образом, что из мезенхимных клеток, прилежащих к глазному бокалу, образуется сосудистая оболочка, в то время как из наружных слоев выделяется фиброзная оболочка. Именно из передней части фиброзной оболочки выделяется прозрачная зона, которая превращается в роговицу. Плод в возрасте 6–8 мес содержит в СО кровеносные сосуды, которые находятся в капсуле хрусталика, а также в стекловидном теле. Они постепенно подвергаются инволюции и исчезают, при этом рассасывается мембрана, которая закрывает отверстие зрачка (зрачковая мембрана) [36, 230, 269, 425].
В свою очередь, верхние и нижние веки, которые идентифицируются с 3-го месяца внутриутробной жизни, вначале имеют вид складок эктодермы. Покровные эпителиоциты конъюнктивы, в том числе и покрывающие роговицу на переднем полюсе, имеют эктодермальное происхождение. Glandula lacrimalis развивается за счёт выростов конъюнктивального эпителия, которые появляются на 3-м месяце пренатального
25

ГЛАВА I
онтогенеза, располагаясь в латеральной части формирующегося верхнего века [35, 50].
Глазное яблоко новорожденного характеризуется относительно большими размерами сравнительно с лицевым черепом, при этом переднезадний размер достигает 17,5 мм, а масса соответствует 2,3 г. Известно, что зрительная ось глазного яблока новорождённого проходит гораздо латеральнее, чем у взрослого человека. При этом глазное яблоко на первом году жизни ребенка растет быстрее, чем во все последующие годы. Только к 5 годам масса глазного яблока увеличивается примерно на 72%, а к 22–27 годам достигает размеров, превышающих в 3 раза таковые у новорожденных [48, 65, 85].
Анатомически роговица у новорожденного человека относительно толстая, при этом кривизна ее в течение всей жизни практически не меняется; а хрусталик имеет почти круглую форму, при этом радиусы его передней и задней кривизны почти равны. Особенно стремительно хрусталик растет в течение только 1-го года жизни, а в дальнейшем темпы роста его значительно снижаются. Радужка характеризуется в этот период развития тем, что имеет выпуклую поверхность на переднем полюсе, пигмента в ней мало по сравнению с последующими периодами развития, диаметр зрачка достигает 2,5 мм. По мере накопления возрастных изменений ребенка толщина iris увеличивается, концентрация меланина в ней возрастает, а диаметр зрачка увеличивается. Многие авторы отмечают, что в возрасте 41–55 лет зрачок незначительно уменьшается [91, 156, 195].
Corporis ciliaris у новорожденного характеризуется слабым развитием. Рост, дифференцировка и специализация ресничной мышцы характеризуются высокой скоростью. Зрительный нерв у новорожденного короткий, имеет незначительную толщину, достигающую 0,8 мм. Но
к25 годам жизни диаметр его возрастает примерно вдвое [228, 294]. Мышцы глазного яблока у новорожденного приобретают доста-
точно хорошее развитие, исключая их сухожильную часть. С этим связано то, что движение глаза может осуществляться сразу после рождения, однако характеризуется тем, что координация этих движений наступает только начиная со 2-го месяца постнатальной жизни ребенка [66, 373].
Glandula lacrimalis новорожденного в первый год жизни имеет относительно небольшие размеры, её выводные канальцы очень тонкие. Слезоотделительная и секреторная функции появляются на 2-м месяце жизни. Влагалище глазного яблока у новорожденных и детей грудного возраста в первые месяцы жизни тонкое, а жировое тело глазницы развито относительно слабо. Выявлено, что у людей пожилого и старческого возраста жировое тело в дистальном отделе глазницы уменьшается
26

СОВРЕМЕННОЕ СОСТОЯНИЕ ВОПРОСА ХОРИОИДАЛЬНОГО АНГИОГЕНЕЗА
в размерах, частично атрофируется, и глазное яблоко начинает меньше выступать из глазницы [109, 249, 409].
Сравнительный анализ показал, что, как правило, глазная щель
убольшинства новорожденных узкая, а её медиальный угол имеет закругленную форму. Многими исследователями отмечено, что в дальнейшем глазная щель имеет тенденцию к быстрому увеличению. При этом
удетей до 15–17 лет она имеет большую ширину, с этим связано то, что глаз кажется относительно больших размеров, чем у взрослого человека [86, 134, 207, 346].
1.4. СТРУКТУРЫ СОСУДИСТОЙ ОБОЛОЧКИ ГЛАЗА ЧЕЛОВЕКА
Большая часть сосудистого тракта или сосудистой оболочки состоит из кровеносных сосудов, питающих внутренние ткани глазного яблока и его содержимое. Наполнение сосудистой оболочки кровью происходит
восновном из четырех задних коротких цилиарных артерий, прободающих склеру обычно в двух местах – чуть кнаружи и внутри от зрительного нерва – и образующих внутри глазного яблока височную и носовую хороидальные ветви (иногда имеется еще и верхняя веточка). Сосудистая оболочка участвует
вподдержании достаточно-
го тургора глазного ябло- |
|
|
ка и обеспечивает тонкие |
|
|
фотохимические процессы |
|
|
в сетчатой оболочке. Из- |
|
|
нутри сосудистая оболочка |
|
|
выстлана пигментным эпи- |
|
|
телием, который принадле- |
|
|
жит уже слоям сетчатки. От |
Рис. 2. Схема строения глаза человека: |
|
количества содержащегося |
||
1 – склера, 2 – сосудистая оболочка, 3 – канал |
||
в этом слое пигмента зави- |
||
шлемма, 4 – корень радужки, 5 – роговица, |
||
сит окраска глазного дна |
||
6 – радужка, 7 –зрачок, 8 – передняя камера |
||
(рис. 2) [8, 21, 73, 87]. |
||
глаза, 9 – задняя камера глаза, 10 – цилиарное |
||
Сосудистая оболочка |
тело, 11 – хрусталик, 12 – стекловидное тело, |
|
глаза имеет толщину 0,05– |
13 – сетчатка, 14 – зрительный нерв, 15 – зо- |
|
0,08 мм и распадается на три |
нулярные волокна [462] |
27

ГЛАВА I
топографически, структурно и функционально различающихся отдела [21, 65]:
1. Передняя часть – радужная оболочка (ирис), окружает центральное отверстие (зрачок) в качестве световой диафрагмы.
Снаружи хориоидея не касается белочной оболочки, так как между ними располагается щель, заполненная ликвором. Увеальному тракту также принадлежит доминирующая роль во внутриглазных обменных процессах. Из всех составляющих сосудистого (увеального) тракта только радужка видна через роговицу. Исключение составляет её крайняя периферия – корень радужной оболочки, прикрытый полупрозрачным лимбом. Эта зона видна лишь при гониоскопии. Прямого контакта с наружной оболочкой она не имеет. Располагается радужка во фронтальной плоскости таким образом, что между ней и роговицей остается свободное пространство – передняя камера глаза, заполненная жидким содержимым – камерной влагой [55, 90, 66].
Радужка не образует плоскости, перпендикулярной анатомической оси глаза. Это связано с тем, что зрачковый край несколько отклонен вперед, поэтому оболочка в целом имеет вид усеченного и уплощенного конуса. Толщина радужки в среднем составляет 300 мкм. Роль радужки заключается не только в экранировании света, но также и в образовании и оттоке внутриглазной жидкости, обеспечении постоянства температуры влаги передней камеры глаза за счет изменения просвета сосудов [90, 293].
Радужка имеет вид тонкой, почти округлой пластинки. Горизонтальный диаметр ее 12,5 мм, вертикальный – 12 мм. Круглое отверстие в центре радужки – зрачок, служит для регулирования количества световых лучей, проникающих в глаз, величина его меняется в зависимости от силы светового потока. Средняя величина его 3 мм, наибольшая – 8 мм, наименьшая – 1 мм. Передняя поверхность радужки имеет радиальную исчерченность, что придает ей кружевной рисунок и рельеф. Исчерченность обусловлена радиальным расположением сосудов, вдоль которых ориентирована строма. Щелевидные углубления в строме радужки называются криптами, или лакунами [96, 137]. Параллельно зрачковому краю, отступя на 1,5 мм, расположен зубчатый валик, или брыжжи, где радужка имеет наибольшую толщину – 0,4 мм. Наиболее тонкий участок радужки соответствует ее корню – 0,2 мм. Брыжжи делят радужку на две зоны: внутреннюю – зрачковую и наружную – ресничную. В наружном отделе ресничной зоны заметны контракционные борозды – следствие сокращения и расправления радужки при ее движении [153, 229].
В радужке различают передний – мезодермальный и задний – эктодермальный, или нейральный (ретинальный), отделы. Мезодермаль-
28

СОВРЕМЕННОЕ СОСТОЯНИЕ ВОПРОСА ХОРИОИДАЛЬНОГО АНГИОГЕНЕЗА
ный листок включает наружный пограничный слой и строму радужки. Эктодермальный листок представлен дилататором с его внутренним пограничным и пигментными слоями. Последние образуют у зрачкового края пигментную бахромку, или кайму. К эктодермальному листку принадлежит и сфинктер, сместившийся в строму радужки по ходу ее эмбрионального развития [120, 247].
Цвет радужки зависит от ее пигментного слоя и присутствия в строме крупных многоотростчатых пигментных клеток. Пигмент поглощает лучи света, которые пробиваются внутрь глаза через открытые части склеры и радужную оболочку. Благодаря этому не происходит диффузного освещения сетчатки. Такое освещение мешало бы усреднению яркости и как следствие – четкости изображения, получающегося от тех лучей, которые проходят через зрачок. Иногда пигмент в радужной оболочке скапливается в виде отдельных пятен. У брюнетов пигментных клеток особенно много, у альбиносов их нет совсем [1, 18].
Радужка имеет две мышцы: сфинктер, суживающий зрачок, и дилататор, обусловливающий его расширение. Сфинктер располагается в зрачковой зоне стромы радужки. Дилататор находится в составе внутреннего пигментного листка, в его наружной зоне. В результате взаимодействия двух антагонистов – сфинктера и дилататора – радужная оболочка выполняет роль диафрагмы глаза, регулирующей поток световых лучей [293, 299, 315]. Дилататор представляет собой очень древнюю форму мышечно-эпителиальных клеток, свойственных простейшим кишечнополостным организмам. Он состоит из веретенообразных клеток. Задняя часть каждой клетки, содержащая ядро, насыщена пигментом. Передняя часть содержит контрактильные фибриллы. Поэтому клетки дилататора относят к миоэпителиальным образованиям. Мускульные волокна дилататора, сгущаясь, сходятся, не достигая зрачкового края, на расстоянии 1,3 мм. На этом уровне они встречают мышечную систему сфинктера. Дилататор зрачка, распространяясь в строме по направлению к сфинктеру, образует структуры в виде выроста мышцы, которые называют шпорами Фукса и Мишеля. По направлению к периферии дилататор постепенно исчезает. Сфинктер образует ленту шириной 1 мм, концентричную по отношению к зрачковому отверстию. Эти два отличные один от другого мускульных слоя связаны между собой соединительнотканными волокнами [53, 54, 248]. Их особенности в детском возрасте заключаются в слабой морфологической выраженности и как следствие – слабом функционировании, с этим связана узость зрачка у детей [25, 204].
Морфологические данные, полученные многими морфологами, свидетельствуют о том, что радужка включает 5 слоев: передний эпителий,
29

ГЛАВА I
расположенный на передней поверхности радужки, наружный пограничный (бессосудистый) слой, сосудистый слой, внутренний пограничный слой и задний двухслойный пигментный эпителий. Передний эпителий ссотоит из плоских полигональных клеток. Он является фактическим продолжением эпителия, который покрывает заднюю поверхность роговицы [63, 168]. Некоторые из авторов утверждают, что эндотелиальное покрытие переднего пограничного слоя, отграничивающее последний от влаги передней камеры, не является постоянной структурой, существуя только на первых годах жизни ребенка. В дальнейшем постепенно эндотелиальные клетки замещаются фибробластами [122, 170]. Толщина переднего эпителия составляет 12,5 мкм. Каждая клетка состоит из апикальной – эпителиальной части и базальной – мышечной, которая проникает в строму радужки. Мышечная поверхность эпителия волнистая, его задняя граница трудно различима. Клетки переднего эпителия имеют базальную мембрану [26, 373]. Апикальная порция переднего эпителия соединяется с задним эпителием, однако между ними существует пространство в 20 нм, заполненное микровиллами. Они исходят из обоих эпителиальных слоев. В это пространство проникают реснички, возникающие из цитоплазмы эпителиальных клеток [273, 402]. Между плоскими клетками переднего эпителия остаются многочисленные поры и межклеточные пространства, которые контактируют с канальцеобразными пространствами переднего пограничного слоя [308, 365]. Вдоль апикальной поверхности клетки соединены многочисленными десмосомами.
Вмышечной порции клеток переднего эпителия десмосомы исчезают, здесь видны лишь замковые соединения. Между боковыми поверхностями клеток существуют интерцеллюлярные пространства [366, 405].
Врайоне апикальной порции клеток базальная мембрана отсутствует.
Вапикальной части находится большое количество клеточных органелл (митохондрии, пигментные гранулы, гранулярный эндоплазматический ретикулум, трубочки гладкого эндоплазматического ретикулума; в перинуклеарной области имеют место скопления свободных рибосом, видны миофиламенты) [97, 370].
Сзади радужка покрыта двухслойным пигментированным задним эпителием. Клетки заднего эпителия носят чисто эпителиальный характер. После депигментации они имеют прямоугольную форму [309, 422]. Высота их составляет 36–55 мкм, ширина 16–25 мкм. Цитоплазма содержит большое число круглых или сигароподобных пигментных гранул, которые являются более крупными, чем в стромальных меланоцитах.
Взрачковой области пирамидальная форма клетки более выражена, ее пигментация здесь не меняется, в то время как по направлению к периферии происходит процесс явной депигментации, усиливающийся
30