

141
Синтез тимидилового нуклеотида:
В процессе синтеза тимидилового нуклеотида происходит восстановление рибозы в дезоксирибозу при участии рибонуклеотидредуктазного комплекса (включающего белок тиоредоксин, витамин В12 и НАДФН2), а урацил метилируется в тимин
Авторегуляция синтеза пиримидиновых нуклеотидов
Ключевыми ферментами являются карбамоилфосфатсинтетаза и карбамоиласпартаттрансфераза
Эти аллостерические ферменты угнетаются высокими концентрациями готовых пиримидиновых нуклеотидов.
Многие противоопухолевые препараты являются ингибиторами фермента тимидилатсинтетазы.
Дополнительный путь синтеза пиримидиновых нуклеотидов возможен из готовых оснований.
9.3. Биосинтез ДНК Возможны 2 пути биосинтеза ДНК в организме: репликация, репарация.
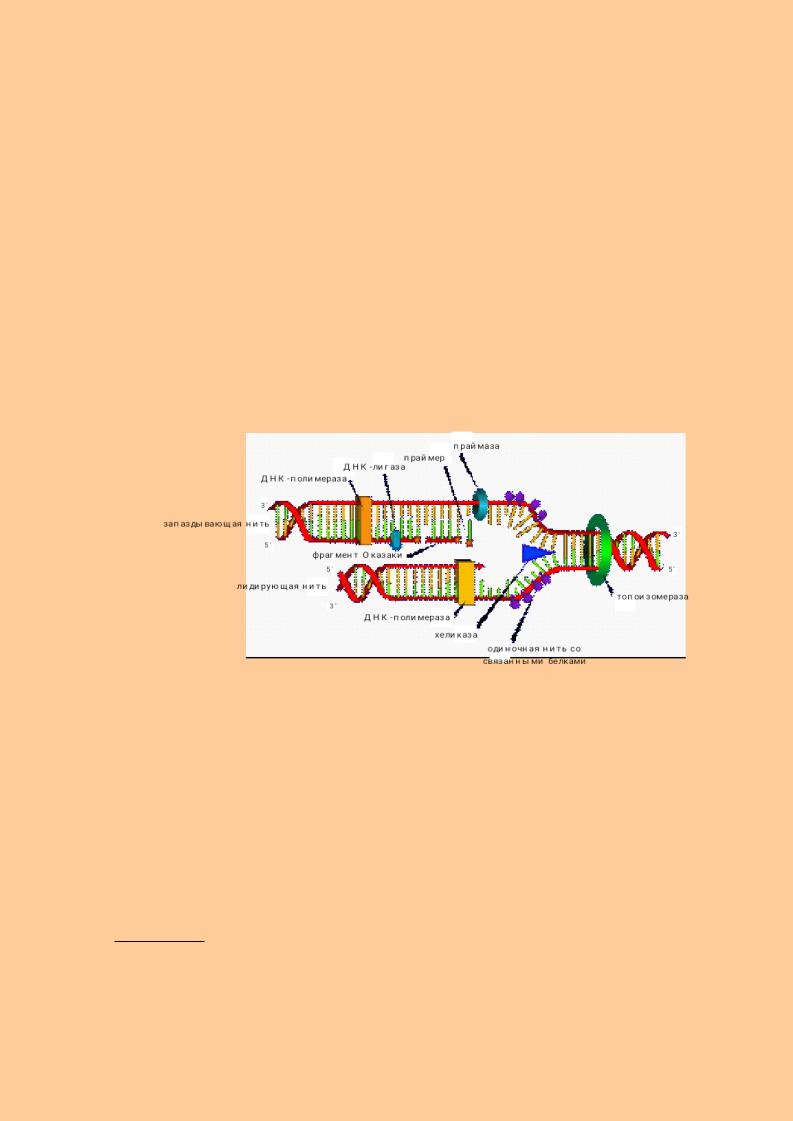
Репликация - самовоспроизведение молекулы ДНК с целью передачи генетической информации. В репликации ДНК участвует сложный репликационный комплекс, который включает в себя:
1.нуклеотиды в виде трифосфатов (АТФ, ГТФ, ТТФ, ЦТФ). Они являются одновременно и структурным материалом и источником энергии;
2.ДНК-матрица – программа, по которой будет синтезироваться ДНК;
3.праймер (затравка) - короткая молекула РНК, к которой присоединяется первый нуклеотид ДНК;
4.ферменты:
142
∙ДНК-полимеразы соединяют нуклеотиды в полинуклеотидную цепь;
∙ДНК-рестриктазы разрывают полинуклеатидную цепь ДНК;
∙ДНК-лигазы соединяют фрагменты ДНК;
∙хеликазы раскручивают молекулу ДНК;
∙топоизомеразы осуществляют кратковременные разрывы ДНК в процессе синтеза и контролируют конформацию вновь синтезированной ДНК;
∙праймазы – осуществляют синтез затравки.
5.белковые факторы;
6.ионы металлов.
Репликация ДНК происходит в S-фазу клеточного цикла. Важную роль в
чередовании циклов выполняют белки - циклины.
Сущность репликации сводится к соединению нуклеотидов в комплементарную антипараллельную дочернюю цепь ДНК. В репликации выделяют 3 стадии: инициация (начало синтеза), элонгация (удлинение цепи), терминация (окончание синтеза).
Матрицей для синтеза ДНК являются обе полинуклеотидные цепи материнской ДНК. Репликация начинается в нескольких участках материнской ДНК (точки репликации, точки Ориджины). В них происходит частичный разрыв водородных связей с формированием репликационных утолщений (узелков). В последующем, при участии ферментов ДНК-рестриктаз происходит разрезание молекулы ДНК. Под действием ферментов хеликаз возникает расплетание (раскручивание) двух цепей, и при участии топоизомераз формируется репликационная вилка, в которой одна цепь имеет направление 5→3, а вторая 3→5. Затем на каждой из цепей синтезируются дочерние нити ДНК по принципу комплементарности. Поскольку ДНКполимераза замыкает связь только в направлении 5’→3’, то на одной цепи ДНК происходит синтез непрерывной (лидирующей) нити ДНК в направлении 5→3. При синтезе лидирующей цепи праймазы синтезируют праймер, затем ДНКполимераза присоединяет к праймеру первый ДНК-овый нуклеотид и по принципу комплементарности происходит удлинение молекулы ДНК. На второй материнской нити ДНК синхронно синтезируется отстающая нить ДНК, которая синтезируется в виде небольших фрагментов в направлении 5→3. В последующем эти фрагменты

143
(фрагменты Оказаки) соединяются между собой ДНК-лигазами. Праймеры при этом расщепляются.
Таким образом, биосинтез ДНК проходит по полуконсервативному типу, при котором в новой ДНК одна цепь материнская, а другая - дочерняя.
Репарация ДНК происходит при появлении в её молекуле повреждений, искажений. При репарации вначале распознаётся место повреждения, затем ферменты рестриктазы вырезают дефектный участок, ДНК-полимеразы по принципу комплементарности синтезируют отсутствующий участок, а ДНК-лигазы прикрепляют его к сохранившимся участкам неповреждённой ДНК.
9.4. Биосинтез РНК (транскрипция)
Транскрипция – синтез молекулы РНК по матрице ДНК. Биологическая роль: перенос генетической информации с ДНК на РНК. В транскрипции участвуют:
∙матрица (программа) – кодирующая нить ДНК;
∙субстраты – АТФ, ГТФ, УТФ, ЦТФ;
∙ферменты – РНК-полимеразы;
∙белковые факторы;
∙ионы магния, марганца.
Выделяют 3 стадии транскрипции: инициация, элонгация, терминация На молекуле ДНК имеется особый участок промотор, с которым
связывается РНК-полимераза. Промотор иногда называют ТАТА участком (в нём преобладают тимин, аденин, с 2-мя водородными связями между ними). Рядом с промотором расположены сигнальные участки, определяющие скорость транскрипции. Далее в молекуле ДНК располагаются кодирующие (экзоны) и некодирующие (интроны) участки гена. Участок (сайт) терминации определяет окончание синтеза РНК.
Инициация заключается во взаимодействии иницирующих белков с промотором и расхождении нитей ДНК, их раскручивании и формировании транскрипционной вилки. РНК-полимераза связывается с промоторным участком и по принципу комплементарности соединяет нуклеотиды цепи РНК в направлении от 5 конца к 3. РНК-полимераза - олигомерный фермент, состоящий из нескольких субъединиц, не требующий затравки. При достижении РНК – полимеразой участка терминации происходит его связывание с белками терминации, что сопровождается отщеплением РНК-полимеразы от ДНК, диссоциацией её и окончанием транскрипции.
Процессинг РНК
Синтезированная РНК переписывает с кодирующей нити ДНК, как кодирующие участки, так и некодирующие участки гена и является про-РНК (незрелой РНК). Про-РНК в последующем подвергается созреванию (процессингу). Существует несколько механизмов процессинга:
∙сплайсинг – вырезание копий интронов и соединение копий экзонов;
∙присоединение к про-РНК добавочных нуклеотидов;
∙модификация азотистых оснований в составе про-РНК.
Особенности процессинга для рРНК,тРНК,иРНК.
Процессинг иРНК заключается в присоединении КЭП - участка и

144
полиаденилового «хвоста» в сочетании со сплайсингом.
Процессинтг тРНК происходит путём метилирования азотистых оснований и добавления акцепторного участка ЦЦА в сочетании со сплайсингом.
Процессинтг рРНК заключается в вырезании из большого предшественника фрагментов всех видов РНК: 18S; 5S; 5,8S; 28S.
Возможен альтернативный сплайсинг, который состоит в том, что для различных видов белков интроны могут служить экзонами.
Возможен и безматричный синтез РНК. Он происходит из нуклеозиддифосфатов при участии фермента полинуклеотидфосфорилазы. Данным способом синтезируются стандартные, небольшие молекулы РНК, необходимые для синтеза стандартных белков.
Таким образом, передача генетической информации происходит в следующем направлении: ДНК→ РНК→ белок. Однако, в некоторых фагах, эмбриональных тканях возможен синтез ДНК по матрице РНК (РНК→ДНК). Этот вариант синтеза катализирует фермент РНК-зависимая ДНК-полимераза (обратная транскриптаза, ревертаза). В вирусах возможен также вариант синтеза РНК→РНК при участии РНК-репликазы.
9.5. Биосинтез белков (трансляция)
Основной структурой синтезируемых белков является первичная структура (последовательность аминокислот в полипептидной цепи), которая заложена в генетическом коде ДНК.
9.5.1. Характеристика генетического кода Генетический код имеет ряд характеристик.
1.Триплетность –1 аминокислота кодируется тремя нуклеотидами. Из 4 видов нуклеотидов ДНК при триплетности кода возможно 64 различных сочетания, что достаточно для кодирования 20 аминокислот.
2.Однозначность – 1 триплет кодирует только 1 аминокислоту.
3.Вырожденность – для кодирования 1 аминокислоты может быть несколько триплетов
4.Непрерывность – между триплетами отсутствуют нуклеотиды, не принадлежащие соседним триплетам.
5.Неперекрываемость – один нуклеотид не может одновременно принадлежать 2-м триплетам.
6.Универсальность – код в разных организмах одинаков, отвечает за одни и
те же аминокислоты.
Таким образом, код ДНК является линейным непрерывным и однонаправленным. Последовательность нуклеотидов строго соответствует последовательности аминокислот в синтезируемом белке – принцип коллинеарности.
9.5.2. Трансляция Для трансляции необходимы следующие факторы:
∙все виды РНК (тРНК, иРНК, рРНК);
∙аминокислоты в активной форме;

145
∙макроэрги;
∙ферменты;
∙добавочные белковые факторы;
∙ионы Mg2+.
На первой подготовительной стадии происходит активация аминокислот и связывание их со «своей» транспортной РНК. В этой стадии участвуют ферменты аминоацил-тРНК-синтетазы. Это специфичные ферменты, обеспечивающие соединение аминокислоты с соответствующей тРНК.
Инициация синтеза белка происходит при образовании инициирующего комплекса, который включает в себя инициирующий кодон (АУГ, АГУ) иРНК, аминоацил - тРНК, рибосому. Информационная РНК своим КЭП-участком соединяется с малой субъединицей рибосомы. К инициирующему кодону присоединяется тРНК с первой аминокислотой (чаще всего метионином). К малой субъединице присоединяется большая субъединица рибосомы, и на рибосоме формируется два функциональных участка: пептидильный (Р-участок)
иаминоацильный (А-участок). Первая тРНК с первой аминокислотой
присоединяется к Р-участку, а А-участок оказывается свободным.
Инициация Элонгация Элонгация включает в себя замыкание пептидной связи, транслокацию
рибосомы по иРНК с использованием энергии ГТФ и АТФ. К свободному А-участку присоединяется своим антикодоном вторая тРНК со второй аминокислотой. Под действием фермента пептидилтрансферазы первая аминокислота отрывается от первой тРНК и присоединяется ко второй аминокислоте с формированием дипептида. В последующем происходит смещение (транслокация) рибосомы по иРНК на расстояние трёх нуклеотидов. При этом вторая тРНК с дипептидом оказывается в пептидильном участке, а аминоацильный участок освобождается. Первая тРНК перемещается из рибосом в цитозоль для соединения с новой аминокислотой, а к А-участку присоединяется третья тРНК с третьей аминокислотой. Затем дипептид переносится на третью аминокислоту сообразованием трипептида. Синтез полипептидной цепи белка осуществляется в направлении от N-конца к С-концу. В процессе трансляции тРНК выполняет своеобразную адапторную роль в переводе четырёхзначной информации иРНК в двадцатизначную информацию в белках.

146
Элонгация |
Транслокация |
|
Терминация происходит |
при приближении белоксинтезирующего |
|
|
|
|
комплекса к терминирующему кодону иРНК (УАГ, УГА). Этому кодону не соответствует ни одна из тРНК, поэтому не приносится новая аминокислота, и синтез белка обрывается.
9.5.3. Посттрансляционная модификация белков Многие синтезированные белки в последующем подвергаются
посттрансляционной модификации. Существует несколько её вариантов. Наиболее часто встречается:
1.частичный протеолиз – отщепление «ненужных» участков (профермент→фермент; прогормон→гормон);
2.модификация отдельных аминокислот:
∙окисление (пролин→гидроксипролин в коллагене);
∙фосфолирирование (присоединение фосфата);
∙гликозилирование (присоединение углевода);
∙карбоксилирование (присоединение группы СООН в некоторых факторах свёртывания крови);
3.присоединение простетической группы;
4.замыкание дисульфидных мостиков;
5.изменение олигомерности белка (объединение нескольких мономеров).
Впосттрансляционной модификации белков играют важную роль белки - шапероны (они «следят» за правильностью модификации).
9.5.4.Ингибиторы биосинтеза белка
Вклинической практике применяют в качестве антибактериальных препаратов ингибиторы синтеза нуклеиновых кислот и белка в микроорганизмах. Они проявляют эффект на различных стадиях:
∙ на стадии репликации Антимицин Д – встраивается в молекулу ДНК и блокирует синтез новой ДНК;
Новобиацин – ингибирует ДНК-гиразу (топоизомеразу); ∙ на стадии транскрипции Рифамицин – блокирует РНК-полимеразу; ∙ на стадии трансляции
Тетрациклин, левомицетин – связывают либо малую, либо большую субъединицу рибосомы и тем самым блокируют синтез белка;
147
Пенициллин – блокирует синтез белков, входящих в оболочку микроорганизмов.
9.5.5. Регуляция биосинтеза белка Наиболее подробно выяснена регуляция синтеза белка на
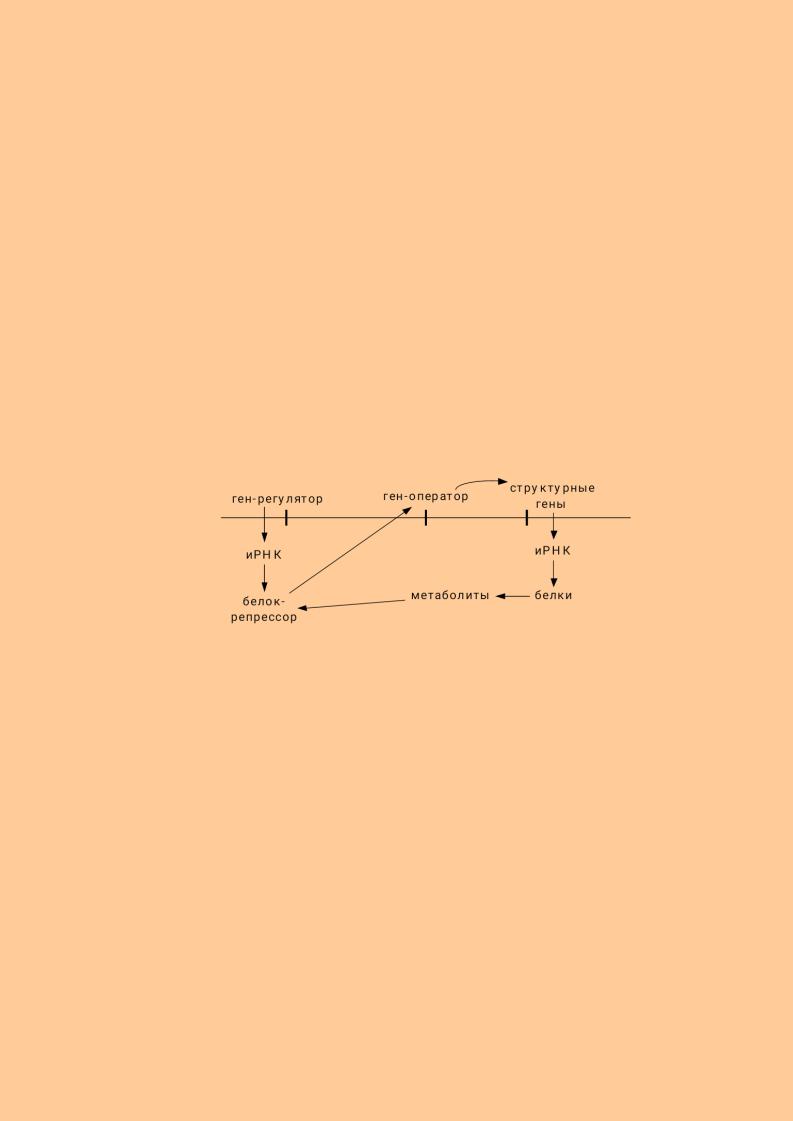
микроорганизмах. Общепринятой является схема авторегуляции Ж. Моно и Ф. Жакоба. Согласно этой схеме, в молекуле ДНК выделяют ген-регулятор, который отвечает за синтез особой РНК и в последующем за синтез особого белка репрессора. На удалении от этого участка находятся ген-оператор и струрктурные гены. Структурные гены (цистроны) служат матрицей для синтеза иРНК, а в последующем – белков. Белки в процессе обмена образуют метаболиты. Метаболиты связываются с репрессором. Репрессор регулирует активность гена – оператора, который, в свою очередь, влияет на структурные гены.
Угнетение биосинтеза белков осуществляется по принципу обратной связи. Высокая концентрация белка и метаболитов приводит к тому, что метаболиты связываются с регрессором и активируют его. Активный репрессор подавляет ген - оператор, а тот - структурные гены, в силу чего синтез белка прекращается.
10. БИОХИМИЯ ГОРМОНОВ Гормоны – органические биологические вещества, вырабатываемые в
эндокринных железах или отдельных клетках, транспортируемые кровью и оказывающие регуляторное действие на обменные процессы и физиологические функции.
Гормоны являются первичными посредниками между центральной нервной системой и тканевыми процессами. К эндокринным железам относится гипоталамус, гипофиз, эпифиз, тимус, щитовидная железа, паращитовидная железа, поджелудочная железа, надпочечники, половые железы и диффузная нейроэндокринная система. Единый принцип номенклатуры гормонов отсутствует. Их называют по месту образования (инсулин от insula-островок), по физиологическому эффекту (вазопрессин), гормоны передней доли гипофиза имеют окончание – тропин, окончание – либерин и – статин указывает на гипоталамические гормоны.
10.1. Классификация гормонов по их химической природе По химической природе гормоны делят на 3 группы.
I.Белково-пептидные гормоны.

148
a)Простые белки (например, соматотропин, инсулин);
b)Пептиды (например, кортикотропин, меланотропин, кальцитонин);
c)Сложные белки (чаще гликопротеиды – тиреотропин, гонадотропин).
II.Гормоны - производные отдельных аминокислот (например, тироксин, адреналин).
III.Стероидные гормоны (производные холестерина – кортикостероиды, андрогены, эстрогены).
Химическая природа гормонов определяет особенности их метаболизма.
10.2. Общие сведения об обмене гормонов Синтез гормонов. Гормоны белковой природы синтезируются по
принципам трансляции. Гормоны - производные аминокислот синтезируются путём химической модификации аминокислот. Стероидные гормоны образуются путём химической модификации холестерина. Некоторые гормоны синтезируются в активной форме (адреналин), другие синтезируются в виде неактивных предшественников (препроинсулин). Некоторые гормоны могут активироваться за пределами эндокринной железы. Например, тестостерон в предстательной железе переходит в более активный дигидротестостерон. Синтез большинства гормонов регулируется по принципу обратной связи (авторегуляция)
Под действием импульсов ЦНС в гипоталамусе синтезируется либерины (кортиколиберин, тиреолиберин, соматолиберин, пролактолиберин, гонадолиберин), которые активируют функцию передней доли гипофиза, и статины, тормозящие функцию передней доли гипофиза (соматостатин, пролактостатин, меланостатин). Либерины и статины регулируют выработку тропных гормонов передней доли гипофиза. Тропины передней доли гипофиза, в свою очередь, активируют функцию периферических эндокринных желез, которые вырабатывают соответствующие гормоны. Высокая концентрация гормонов тормозит либо выработку тропных гормонов, либо выработку либеринов (отрицательная обратная связь).
При нарушении регуляции синтеза гормонов может возникать либо гиперфункция, либо гипофункция.
Транспорт гормонов. Водорастворимые гормоны (белково-пептидные

149
гормоны, гормоны - производные аминокислот (исключая тироксин)) транспортируются свободно в виде водных растворов. Водонерастворимые (тироксин, стероидные гормоны) транспортируются в комплексе с транспортными белками. Например, кортикостероиды транспортируются белком транскортином, тироксин - тироксинсвязывающим белком. Белковосвязанные формы гормона расцениваются как определённое депо гормонов. Концентрация гормонов в плазме крови очень мала, находится в диапазоне 10-15-10-19 моль.
Циркулирующие в крови гормоны оказывают эффект на определённые ткани – мишени, в которых имеются рецепторы к соответствующим гормонам. Рецепторы чаще всего являются олигомерными гликопротеидами или липопротеидами. Рецепторы к различным гормонам могут располагаться или на поверхности клеток, или внутри клеток. Количество рецепторов, их активность может изменяться под действием различных факторов.
Катаболизм гормонов.
Гормоны белковой природы распадаются до аминокислот, аммиака, мочевины. Гормоны - производные аминокислот инактивируются различными способами – дезаминирование, отщепление йода, окисление, разрыв кольца. Стероидные гормоны инактивируются путём окислительно-восстановительных превращений без разрыва стероидного кольца, путём реакции конъюгирования с серной кислотой и глюкуроновой кислотой.
10.3. Механизмы действия гормонов Различают несколько механизмов реализации гормонального сигнала для
водорастворимых и водонерастворимых гормонов. Все гормоны оказывают три конечных эффекта:
1)изменение количества белков и ферментов за счёт изменения скорости их синтеза;
2)изменение активности имеющихся в клетки ферментов;
3)изменение проницаемости клеточных мембран.
10.3.1. Цитозольный механизм действия гидрофобных (липофильных) гормонов Липофильные гормоны способны проникать в клетку через клеточную мембрану, поэтому рецепторы для них располагаются внутриклеточно в цитозоле, на митохондриях, на поверхности ядра. Рецепторы гормонов чаще всего включают 2 домена: для связывания с гормоном и для связывания с ДНК. Рецептор при взаимодействии с гормоном изменяет свою структуру, освобождается от шаперонов, в результате чего гормон - рецепторный комплекс приобретает способность проникать внутрь ядра и
взаимодействовать с определёнными участками ДНК. Это, в свою очередь, ведёт к изменению скорости транскрипции (синтез РНК), а вследствие этого меняется и скорость трансляции (синтез белка).
150
10.3.2. Мембранный механизм действия водорастворимых гормонов Водорастворимые гормоны не способны проникать через
цитоплазматическую мембрану. Рецепторы для данной группы гормонов располагаются на поверхности клеточной мембраны. Поскольку гормоны не проходят внутрь клеток, между ними и внутриклеточными процессами необходим вторичный посредник, который передаёт гормональный сигнал внутрь клетки. В качестве вторичных посредников могут служить инозитолсодержащие фосфолипиды, ионы кальция, циклические нуклеотиды.
10.3.2.1. Циклические нуклеотиды - цАМФ, цГМФ - вторичные посредники Гормон взаимодействует с рецептором и образует гормон - рецепторный
комплекс, в котором меняется конформация рецептора. Это, в свою очередь, изменяет конформацию мембранного ГТФ - зависимого белка (G-белка) и ведёт к активации мембранного фермента аденилатциклазы, который переводит АТФ в цАМФ.
Внутриклеточный циклический АМФ служит вторичным посредником. Он активирует внутриклеточные ферменты протеинкиназы, которые катализируют фосфорилирование различных внутриклеточных белков (ферментов, мембранных белков), что приводит к реализации конечного эффекта гормона. Эффект гормона «выключается» под действием фермента фосфодиэстеразы, разрушающей цАМФ, и ферментов фосфатаз, дефосфорилирующих белки.
.  10.3.2.2. Ионы кальция -вторичные посредники
10.3.2.2. Ионы кальция -вторичные посредники
Взаимодействие гормона с рецептором повышает проницаемость кальциевых каналов клеточной мембраны, и внеклеточный кальций поступает в цитозоль. В клетках ионы Са2+ взаимодействуют с регуляторным белком