

101
7.11.1.2. Окисление глицерина На первом этапе глицерин активируется в глицерофосфат, затем
окисляется до фосфодигидроксиацетона, который через стадию образования пирувата переходит в ацетил – КоА и окисляется в цикле Кребса.
7.11.2. Синтез триацилглицеринов (липогенез)
Для синтеза ТАГ необходимы глицерин и жирные кислоты.
7.11.2.1. Синтез глицерина Если глицерин поступает с пищей в недостаточном количестве, он может
синтезироваться из углеводов через стадию образования общего метаболита фосфодигидроксиацетона.
Активная форма глицерина – глицерофосфат используется на синтез ТАГ и глицерофосфолипидов.
7.11.2.2. Синтез жирных кислот Синтез насыщенных жирных кислот происходит в цитозоле при участии
сложного полиферментного комплекса (синтетаза жирных кислот или пальмататсинтетаза). Этот комплекс включает в себя особый ацилпереносящий белок и 6 ферментов. Для синтеза жирных кислот донором водорода является НАДФН2, образующийся в пентозофосфатном пути окисления глюкозы. Исходным веществом для синтеза жирных кислот является ацетил - КоА, который образуется в митохондриях в результате бета – окисления жирных кислот, и самостоятельно выйти в цитозоль не может. Транспорт ацетил - КоА из митохондрий в цитозоль осуществляется с помощью цитратного челночного механизма:
Ацетил - КоА на первом этапе карбоксилируется в малонил - КоА

102
Затем ацетил - КоА и малонил - КоА соединяются с полиферментным комплексом (синтетаза жирных кислот), в котором имеется две SН-группы, принадлежащие ацилпереносящему белку.
Затем остаток ацетила переносится на остаток малонила с образованием β – кетоацил - полиферментного комплекса и освобождением одной SH – группы полиферментного комплекса.
Впоследующем к этому комплексу с четырёх углеродной масляной кислотой присоединяется новая молекула малонил - КоА и в результате синтезируется капроновая кислота (С6) и далее до С16 (пальмитиновая кислота). Более длинные жирные кислоты синтезируется с участием дополнительных ферментов элонгаз.
Втканях организма человека синтезируется только мононенасыщенные кислоты:
олеиновая и пальмитолеиновая. Они образуются из соответствующих насыщенных жирных кислот при участии ферментов десатураз

103
(монооксигеназ) и цитохрома Р450.
7.11.2.3. Синтез триацилглицеринов ТАГ синтезируются из активной формы глицерина и жирных кислот через
стадию фосфатидной кислоты.
7.12. Обмен глицерофосфолипидов
7.12.1. Синтез глицерофосфолипидов Глицерофосфолипиды синтезируются из фосфатидной кислоты и
добавочных азотсодержащих веществ. В синтезе фосфолипидов активатором служит ЦТФ, который может активировать или фосфатидную кислоту, или серин (коламин, холин). В связи с этим возможны два варианта синтеза фосфолипидов.
а) Активация фосфатидной кислоты:
б) Активация холина (серина, коламина):
Таким образом, синтез глицерофосфолипидов и триацилглицеринов на начальных стадиях осуществляется путём предшествующего образования фосфатидной кислоты. Для активации синтеза из неё структурных фосфолипидов используются липотропные вещества. К ним относится холин, метионин, витамин В12 и др. Липотропные вещества препятствуют жировой дистрофии печени, миокарда при гепатитах, миокардитах.
7.12.2. Распад глицерофосфолипидов Распад фосфолипидов осуществляется тканевыми фосфолипазами.
Различают несколько видов фосфолипаз (фосфолипазы А1, А2, С, D), которые разрывают различные связи в фосфолипидах.

104
При ряде патологических состояний наблюдается активация тканевой фосфолипазы А2. Это сопровождается образованием продуктов неполного распада фосфолипидов - лизофосфолипидов. Накопление лизофосфолипидов в составе клеточной мембраны повышает её проницаемость для Na+, К+, Н2О, в результате чего возможен лизис клеток (в частности, гемолиз эритроцитов). Фосфолипаза А2 присутствует в яде некоторых змей.
7.13. Обмен сфинголипидов
7.13.1. Синтез сфинголипидов Сфингозин является компонентом всех сфинголипидов.
Синтез сфингозина происходит из активной формы пальмитиновой кислоты и аминокислоты серина.
Структурным блоком сфинголипидов является церамид, который включает в свой состав сфингозин и жирную кислоту, присоединённую амидной связью.
Синтез церамида осуществляется из активной формы жирной кислоты и сфингозина.

105
Синтез сфингофосфолипидов происходит из церамида и активной формы холина.
ЦДФ – холин (цитидиндифосфохолин) + церамид ( сфингофосфолипид (сфингомиелин)
Синтез цереброзидов осуществляется из церамида и активной формы моносахаров.
Церамид + УДФ - галактоза (уридиндифосфатгалактоза) (цереброзид Синтез ганглиозидов происходит путём присоединения к церамиду
олигосахаридного фрагмента, на конце которого чаще всего находятся сиаловые кислоты и аминогексозы.
7.13. 2. Распад сфинголипидов Расщепление сфинголипидов происходит при участии лизосомальных
ферментов. При их дефекте развивается сфинголипидозы, при которых в ткани головного мозга накапливается сфинголипиды и продукты их неполного распада. Сфинголипидозы проявляются неврологическими симптомами. Для их антенатальной диагностики используется определение содержания сфинголипидов в амниотической жидкости.
Распад сфингофосфолипидов (сфингомиелинов) осуществляется неспецифичными эстеразами и специфичным ферментом сфингомиелиназой, который расщепляет в сфигофосфолипидах связь между церамидом и фосфохолином. При отсутствии данного фермента развивается сфингомиелиноз (болезнь Нимана-Пика).
Распад цереброзидов происходит под действием бета - гликозидазы, расщепляющей связь между церамидом и моносахаридом. При отсутствии данного фермента развивается цереброзидоз (болезнь Гоше)
Распад ганглиозидов осуществляется различными гликозидами, включая гексаминидазу. При отсутствии данного фермента развивается ганглиозидоз (болезнь Тея - Сакса)
7.14. Обмен холестерина Ежесуточно в организме взрослого человека обменивается до 1,5 г
холестерина. Примерно половина этого количества синтезируется в организме (эндогенный холестерин), а половина поступает с пищей (экзогенный холестерин). Холестерин пищевых продуктов всасывается в тонком кишечнике при участии жёлчных кислот.
7.14.1. Биосинтез холестерина Синтез холестерина происходит в печени из ацетил-КоА. Синтез
холестерина представляет собой сложный многоэтапный процесс, протекающий в 20 стадий. Начальная стадия – образование мевалоновой кислоты является

106
ключевой ГМГ - редуктаза – ключевой фермент синтеза холестерина, она угнетается
высокими концентрациями холестерина. Синтезированный в печени холестерин включается в состав липопротеидов ЛПОНП. Под действием липопротеидной липазы ЛПОНП переходят в ЛПНП, которые транспортируют холестерин от печени к органам и тканям. В тканях имеются рецепторы к липопротеидам, при участии которых происходит захват холестерина и проникновение его в клетки.
В клетках часть холестерина превращается в эфиры при участии фермента АХАТ (ацилхолестеролацилтрансфераза). Эфиры холестерина депонируются в тканях.
7.14.2. Использование холестерина в тканях Холестерин необходим для всех клеток и тканей.
1. В печени примерно половина синтезирующегося холестерина превращается в жёлчные кислоты при участии ключевого фермента 7-α-гидроксилазы. Применение веществ, адсорбирующих жёлчные кислоты в кишечнике, повышает переход холестерина в жёлчные кислоты
и снижает его уровень в крови.
2.Холестерин используется для построения клеточных мембран, где он составляет примерно треть всех липидов мембран и определяет физико-химические свойства липидной фазы мембран.
3.В надпочечниках, половых железах холестерин используется на синтез стероидных гормонов
4.В коже из производного холестерина происходит образование витамина Д3 (холекальциферола).
7.14.3. Выведение холестерина из организма Избыток холестерина удаляется из тканей при участии ЛПВП, которые
адсорбируют холестерин из клеток и переносят его в печень. Основная часть холестерина выводится из организма через кишечник в виде жёлчных кислот, продуктов их обмена и образующихся из холестерина под действием микрофлоры холестанола и копростанола. Выведение холестерина из

107
организма в небольших количествах происходит со слущивающимся эпителием, с мочой в виде соединений стероидных гормонов с глюкуроновой кислотой.
7.14.4. Нарушение обмена холестерина
Внорме концентрация холестерина в крови взрослых людей составляет 3,5
–5,2 ммоль/л. У детей концентрация холестерина в крови ниже, чем у взрослых. У новорожденных уровень холестерина равен 2,67 ммоль/л, у детей в возрасте одного года - 4,03 ммоль/л.
Симптом повышения уровня холестерина в крови называется гиперхолестеринемией. Врождённые гиперхолестеринемии встречаются редко, чаще развиваются приобретённые (вторичные) гиперхолестеринемии. На фоне гиперхолестеринемии возможно развитие таких заболеваний как атеросклероз и жёлчекаменная болезнь.
При атеросклерозе избыток холестерина откладывается в эндотелии сосудов, что ведёт к развитию асептического воспаления, отложению кальция, вследствие чего нарушается кровоснабжение тканей. Для диагностики атеросклероза рекомендуется определение коэффициента атерогенности, который показывают соотношение между ЛПНП и ЛПВП.
К атерогенности = (Хобщ. - ХЛПВП)/ХЛПВП ≤ 3.
Для лечения атеросклероза применяют ингибиторы ГМГ-редуктазы, которые блокируют синтез холестерина.
Жёлчекаменная болезнь связана с нарушением соотношения между водонерастворимым холестерином и гидрофильными фосфолипидами и жёлчными кислотами в составе жёлчи. Холестерин является основой формирования камней в жёлчных путях.
При циррозах печени, гепатитах возможно развитие гипохолестеринемии.
7.15. Взаимосвязь липидного и углеводного обменов Обмен углеводов и липидов тесно взаимосвязан как в физиологических
условиях, так и при патологии. Эта взаимосвязь возможна благодаря наличию общих метаболитов в обмене углеводов и липидов.
Углеводы могут использоваться для синтеза различных классов липидов. Некоторые возможные пути перехода углеводов в липиды:
1.Глюкоза→фосфодигидроксиацетон→глицерофосфат→ТАГ, ФЛ
2.Глюкоза→ацетил-КоА→жирные кислоты, холестерин→липиды
3.Глюкоза→ ацетил-КоА→ холестерин→ стероиды
4.Глюкоза→НАДФН2(пентозофосфатный путь)→синтез жирных кислот, синтез сфингозина, синтез холестерина.
Удетей углеводы очень активно используются для синтеза липидов.
Схема взаимодействия углеводного и липидного обмена.
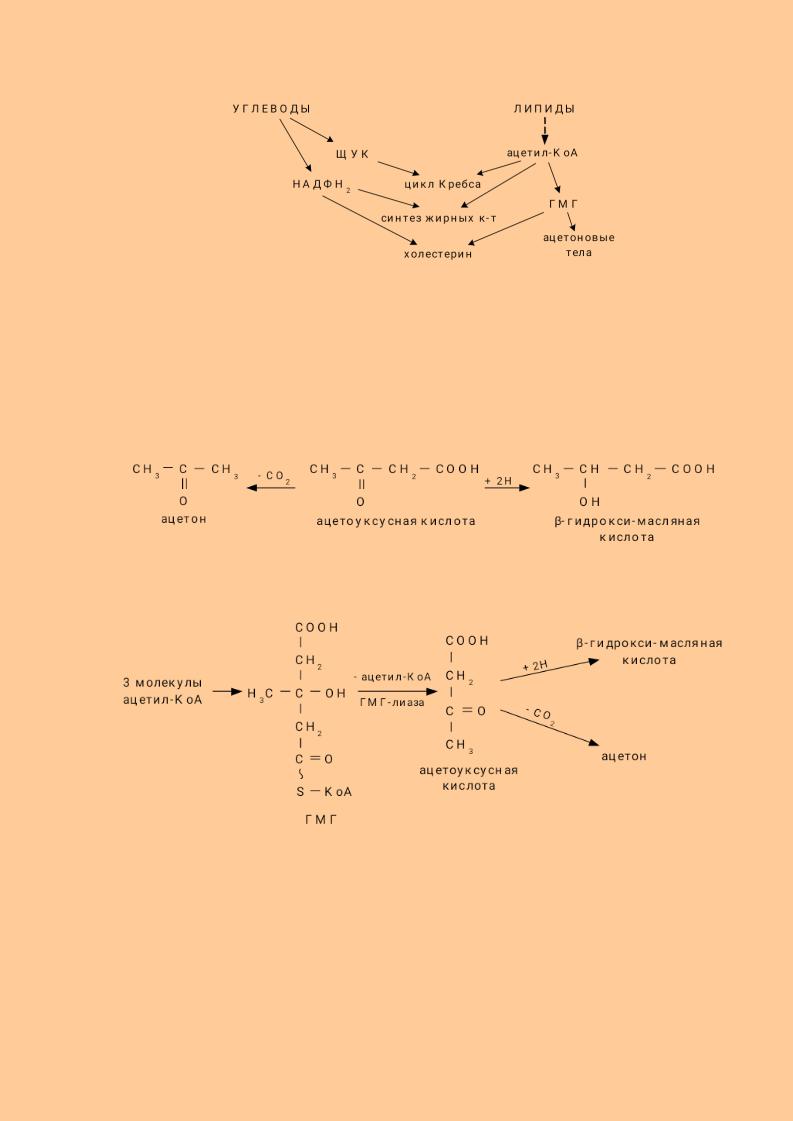
108
У детей углеводы очень активно используются для синтеза липидов.
7.15.1. Ацетоновые тела Распад липидов ведёт к образованию веществ, называемых ацетоновыми
телами. К ацетоновым телам относятся:
Основная масса ацетоновых тел синтезируются в печени из ацетил - КоА через стадию гидроксиметилглютарил - КоА (ГМГ) по схеме:
Внорме концентрация ацетоновых тел очень мала, составляя не более 0,6 ммоль/л. В физиологических условиях ацетоновые тела являются важным водорастворимым энергетическим материалом для различных тканей. Из
печени они транспортируются в ткани, где активируются под действием НS-КоА или сукцинил–КоА (ацетоуксусная кислота + HS-КоА→ацетоацетил - КоА).
Активная форма ацетоновых тел окисляется в цикле Кребса с образованием энергии (ацетоацетил - КоА→24 АТФ).
Впатологических условиях при высокой концентрации ацетоновых тел развивается кетоацидоз. Основными причинами кетоацидоза являются длительное углеводное голодание и сахарный диабет. У детей выражена

109
склонность к кетоацидозу, поскольку у них ограничены запасы гликогена как энергетического материала. В детском возрасте выше скорость распада ТАГ, так как очень лябильна ТАГ-липаза. У детей активно протекает распад кетогенных аминокислот. В то же время усвоение ацетоновых тел в детском возрасте снижено.
7.16. Регуляция липидного обмена На состояние липидного обмена влияют многие физиологические факторы,
в том числе калорийность рациона, интенсивность физической нагрузки. Нервная регуляция липидного обмена подтверждается тем, что жировая
ткань богато иннервирована.
Эндокринная регуляция осуществляется гормонами гипофиза (липотропин, соматостатин), щитовидной железы (тироксин), надпочечников (адреналин, глюкокортикоиды), поджелудочной железы (инсулин, глюкагон), половых желез (андрогены и эстрогены).
Инсулин активирует синтез ТАГ из глюкозы (липогенез) за счёт активации фермента ацетил - КоА – карбоксилазы. Одновременно инсулин обладает антилиполитическим действием (тормозит липолиз) за счёт активирующего влияния на ключевой фермент – ТАГ - липазу.
Большинство других гормонов, участвующих в регуляции липидного обмена, угнетают липогенез, и в большей степени активирует липолиз. Так, гормоны адреналин и глюкагон активируют ТАГ – липазу, а глюкокортикоиды индуцируют синтез ТАГ - липазы.
В регуляции обмена холестерина участвует гормоны тироксин и эстрогены, которые снижают уровень холестерина в крови.
Существует авторегуляция отдельных звеньев липидного обмена. Например, синтез холестерина регулируется по принципу обратной связи (высокая концентрация холестерина в клетках угнетает фермент ГМГ - редуктазу и, тем самым, уменьшает его образование)
7.17. Патология липидного обмена Нарушения обмена липидов возможны на этапе их переваривания,
транспорта, тканевого обмена.
Переваривание липидов нарушается при заболеваниях поджелудочной железы (недостаток ферментов), печени и жёлчевыводящих путей (отсутствуют жёлчные кислоты, страдает эмульгирование, всасывание). При нарушении переваривания и всасывания липидов развиваются авитаминозы жирорастворимых витаминов, истощаются жировые депо, возникает дефицит липидов в организме как основного резервного энергетического материала. Наблюдается потеря липидов через кишечник – стеаторея.
Нарушение транспорта липидов связано с нарушением обмена липопротеидов крови. Примерами врождённых нарушений являются β-алипопротеинемия (болезнь Танжера), α-алипопротеинемия, семейная гиперхолестеринемия. Гораздо чаще встречается приобретённые дислипопротеинемии, при которых изменено соотношение между липидами в различных липопротеидах, в частности, гиперлипопротеинемии – повышение

110
уровня каких либо видов липидов крови.
Нарушение тканевого обмена липидов часто сочетается с нарушением углеводного обмена (голодание, ожирение, сахарный диабет), что может вести к накоплению в тканях, крови ацетоновых тел.
При голодании снижается выработка инсулина, активируется образование глюкагона и адреналина. В результате усиливается распад липидов в тканях и их окисление в качестве основного энергетического материала. Распад липидов приводит к образованию большого количества ацетил - КоА, который при голодании не может быть использован на синтез жирных кислот, холестерина, не может окисляться в цикле Кребса (в силу дефицита щавелевоуксусной кислоты и НАДФН2). Избыток ацетил - КоА используется на синтез ацетоновых тел, поэтому длительное голодание сопровождается выделением ацетоновых тел с мочой.
При сахарном диабете дефицит инсулина сопровождается нарушением усвоения глюкозы тканями, активацией липолиза, образования ацетил - КоА и, как следствие, повышенным образованием ацетоновых тел.
Ожирение может возникать в силу нарушения характера питания (алиментарное ожирение), при эндокринных заболеваниях, длительном применении некоторых лекарственных препаратов. Возможен генетический вариант ожирения, связанный со сбоями в работе гена ожирения, который регулирует синтез гормонов лептинов, активирующих липолиз. При алиментарном ожирении выражена стадийность изменений обмена веществ в организме. В начале патологического состояния активируется выработка инсулина, а в последующем инсулярный аппарат истощается, и возникает относительное преобладание контринсулярных гормонов глюкокортикоидов, развивается своеобразное состояние стероидного диабета, проявляющегося ожирением и повышенным синтезом ацетоновых тел.
Жировая дистрофия миокарда, печени может развиваться после миокардита, гепатита в силу увеличения отложения ТАГ в миокардиоцитах и гепатоцитах. Для профилактики жирового перерождения тканей показано применение различных липотропных веществ, способствующих синтезу структурных глицерофосфолипидов.
В последние годы накапливаются данные о митохондриальных болезнях, при которых страдает β-окисление жирных кислот и нарушается энергетический обмен в тканях.
7.18. Перекисное окисление липидов (ПОЛ)
ПОЛ – неферментативный свободно-радикальный процесс, в который в основном вовлекаются фосфолипиды клеточных мембран.
Инициатором ПОЛ являются активные радикальные формы кислорода: супер - оксид О2-., гидроксирадикал ОН* (О2* + Н2О2→О2 + ОН + ОН*), оксид азота NО*, и пероксинитрил (NO* + О2*→ОNОО). Активные формы О2 атакуют атомы углерода в ненасыщенных жирных кислотах, находящиеся между двойными связями.
Выделяют стадии инициации, разветвления и обрыва процесса ПОЛ Инициация процесса заключается в том, что активные формы О2 отрывают
атом Н от жирной кислоты, превращая жирную кислоту в радикал жирной