

Ядро эукариотической клетки и организация генома эукариот
Средний диаметр ядра обычной клетки животного организма равен 5 мкм, с объемом, составляющим 65 мкм3. За исключением периода деления клетки, ядро плотно и почти равномерно заполнено ДНК. Вследствие своих кислотных свойств, ДНК окрашивается основными красителями, и в свое время, задолго до возникновения современной биохимии и молекулярной биологии ядерное вещество, окрашивающееся этими красителями, получило название хроматина.
Простейшими эукариотическими организмами можно считать протистов или простейших. Однако при всей сложности их организации это далеко не вершина эволюции эукариот. Гораздо более впечатляющие достижения на эволюционном пути были получены не посредством концентрирования усложнений в одной клетке, а путем разделения обязанностей между различными типами клеток.
Разные типы клеток многоклеточного организма часто выглядят совершенно по-разному, несмотря на то, что все они произошли от одной исходной клетки яйцеклетки. При специализации клеток они сохраняют весь объем генетической информации, а это означает, что в основе специализации лежит не потеря или приобретение генов, а изменение порядка и сочетания их экспрессии. У эукариотических клеток развились очень сложные механизмы контроля экспрессии генов. При дифференцировке клеток координированно изменяется и состав мембран, и цитоскелет, и секретируемые продукты и даже метаболизм. Радикальные различия, наблюдаемые при специализации клеток, обусловлены стабильными изменениями в экспрессии их генов.
В эукариотических клетках содержится большое количество ДНК. В клетках человека ее почти в 1000 раз больше, чем в типичной бактериальной клетке, а в клетках некоторых амфибий в 10 раз больше, чем в клетках человека. Однако, известно, что только незначительная часть ядерной ДНК действительно кодирует белки (в клетках человека эта часть составляет около 1%). Возникает закономерный вопрос зачем нужны остальные 99% ДНК. В настоящее время обсуждаются две гипотезы: одна из них утверждает, что эта ДНК просто увеличивает массу ядра, в соответствии с другим предположением данная ДНК представляет собой собрание уже бесполезных для клетки последовательностей, десятками и сотнями тысячелетий накапливавшихся в ней и обеспечивавших в прошлом функционирование механизмов биосинтеза и размножения.
Действительно, в молекулах ДНК многих видов обнаружены так называемые псевдогены – определенные последовательности, которые в настоящее время не экспрессируются, возможно, вследствие утраты функциональной активности их регуляторных участков. Типичным примером может служить присутствие двух псевдогенов -глобина в 11 хромосоме и одного псевдогена -глобина в 16 хромосоме человека.
С другой стороны в молекулах ДНК присутствуют «подвижные» (мобильные) элементы, представленные соответствующими последовательностями, которые могут внезапно «перепрыгивать» из одного участка ДНК в другой и даже встраивать в новые места молекул ДНК свои дополнительные копии. Таким образом, эти мобильные элементы могут размножаться подобно медленно развивающейся внутренней инфекции, все больше увеличивая объем генетического материала.
Несмотря на огромные достижения в изучении механизмов «работы» геномов, современные знания о строении и функционировании ДНК далеко не совершенны. Поэтому не исключено, что некодирующая ДНК выполняет определенную важную функцию, пока еще неизвестную.
Тем не менее, в настоящее время уже установлено, что часть этой ДНК выполняет структурную функцию, позволяя генетическому материалу конденсироваться и плотно упаковываться в ядерном пространстве. Другая часть «лишней» ДНК регуляторная участвует в процессах, обеспечивающих включение и выключение генов, направляя, тем самым, синтез белков, и играя, таким образом, ключевую роль в сложных механизмах регуляции экспрессии генов эукариотической клетки. Кроме того, определенные участки ДНК, не кодирующие белки, ответственны за синтез рибосомных и транспортных РНК, принимающих непосредственное участие в процессах трансляции информационных РНК.
В процессе эволюции эукариотические клетки должны были решить проблему упаковки столь огромного количества генетического материала в ограниченном объеме клеточного ядра. Действительно, ДНК эукариот имеет настолько большую суммарную длину, что риск ее запутывания или разрыва при манипуляциях с нею in vivo очень велик. Общая длина ядерной ДНК в клетках млекопитающих составляет приблизительно 2 м. Если считать средний объем ядра, равным 65 мкм3, то проблема упаковки всей ДНК может быть сведена к решению следующей задачи: разместить в мяче для игры в теннис 18 км нити, имеющей несжимаемый диаметр в 1 мм .
Индивидуальные эукариотические хромосомы хорошо различимы только в течение короткого промежутка времени – в процессе клеточного деления. Именно в М-фазе клеточного цикла каждая хромосома имеет вид компактного образования с плотностью упаковки ДНК около 10.000. В течение же бóльшей части жизненного цикла эукариотической клетки генетический материал присутствует в ядре в виде интерфазного хроматина – зернистых участков, выявляемых с помощью определенных оснóвных красителей.
Различают два типа хроматина. В бóльшей его части хроматиновые нити упакованы менее плотно, чем в хромосоме. Этот ядерный материал называют эухроматином. Обобщенная плотность его упаковки в интерфазе составляет приблизительно 1000 – 2000. В некоторых участках хроматиновые нити упакованы очень плотно и находятся в состоянии, напоминающем по степени упаковки митотические хромосомы. Этот материал называют гетерохроматином. Между структурой генетического материала и его транскрипционной активностью существует определенная корреляция. Так, митотические хромосомы транскрипционно инертны, т.е. в период клеточного деления процессы транскрипции приостанавливаются. В гетерохроматине транскрипционная активность также отсутствует. Описано два класса гетерохроматина, каждый из которых содержит последовательности разного типа, но они практически в равной степени лишены транскрипционной активности.
Конститутивный гетерохроматин состоит из определенных областей, которые никогда не экспрессируются. К этому классу относятся сателлитные последовательности ДНК. В виде факультативного гетерохроматина могут находится целые хромосомы, которые неактивны в целом ряду клеточных поколений, однако, при определенных обстоятельствах они способны к экспрессии. Примером подобной ситуации служит Х-хромосома млекопитающих, одна из копий которой (случайная) полностью инактивирована у каждой самки. Неактивная Х-хромосома является составной частью гетерохроматина, в то время как активная Х-хромосома входит в состав эухроматина.
Отсутствие активности связано с чрезвычайно высокой степенью конденсации генетического материала. Несмотря на то, что активные гены действительно находятся в неконденсированном эухроматине, лишь незначительная их часть способна к транскрипции. Таким образом, необходимым условием экспрессии генов является их локализация в эухроматине, но одного этого условия явно недостаточно.
Одним из наиболее замечательных достижений молекулярной биологии последних лет явилось установление общей структуры хроматина. До относительно недавнего времени было известно только то, что хроматин содержит ДНК, РНК и белки, организованные в компактную форму, в виде которой бóльшая часть последовательностей ДНК являются структурно недоступными и функционально неактивными. Относительная доля указанных компонентов хроматина может варьировать в зависимости от источника и метода выделения. В целом, масса белка почти в два раза превышает массу ДНК, а масса РНК составляет менее 10% массы ДНК. В свое время полагали, что отсутствие транскрипционной активности в хроматине связано с образованием нуклеопротеидных комплексов, структура которых, однако, не была известна.
Необычайный прогресс был сделан в 1974 году, когда удалось установить, что хроматин сконструирован из частиц, имеющих одинаковый тип организации у всех эукариот.
Если выделенные и отмытые интерфазные ядра ресуспендировать в растворе с низкой ионной силой, они в результате набухания и последующего лизиса высвобождают нити хроматина. Исследование хроматина под электронным микроскопом позволило обнаружить два типа хроматиновых нитей: волокна диаметром 10 нм (100 Å) и волокна диаметром 30 нм (300 Å). Нити размером 30 и 10 нм могут обратимо превращаться друг в друга при изменении ионной силы раствора. При высокой ионной силе обнаруживаются преимущественно волокна диаметром 30 нм.
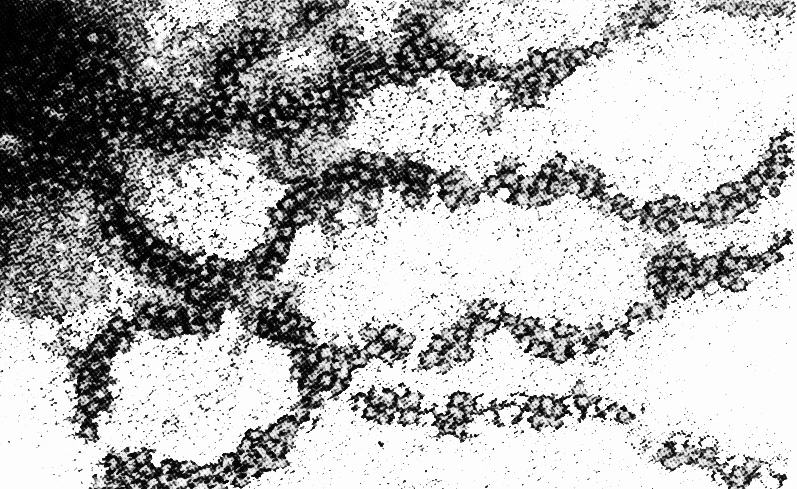
Рис. 2.2 |
Электронная микрофотография нити диаметром 30 нм, которая имеет спиральную структуру (Lewin B., Genes III, New York, Wiley, 1987). |
На электронно-микроскопических фотографиях, хроматиновые волокна, имеющие диаметр 10 нм, часто обнаруживаются в двух альтернативных формах: в форме сплошного волокна (рис. 2.3 А) либо, чаще, в форме более вытянутого волокна с утолщениями – «бусинками», нанизанными по всей длине нити с определенными интервалами (рис. 2.3 В). Данные утолщения представляют собой нуклеопротеидные комлексы, образованные сегментом ДНК определенной длины, связанным с cильнооснóвными белками особого типа – гистонами.
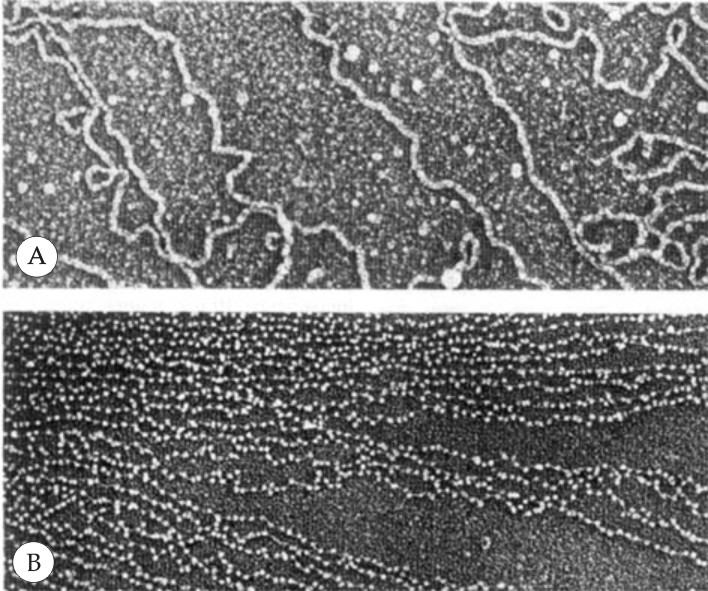
Рис. 2.3 |
Электронные микрофотографии хроматина. А – волокно хроматина диаметром 10 нм из почечных клеток CV1 обезьяны. В – хроматин из эритроцитов цыпленка, имеющий вид нити с нанизанными на нее «бусинками» (Сингер М., Берг П., Гены и геномы, М., «Мир», 1998, т. 1, с. 50). |
Каждая из таких «бусинок» представляет собой организованную структуру, которая получила название нуклеосомы. В особенно вытянутых участках хроматина видно, что индивидуальные нуклеосомы соединены тонкой нитью – это свободная двухцепочечная ДНК. Индивидуальные нуклеосомы можно получить посредством обработки хроматина ДНК-азой микрококков. Данная эндонуклеаза расщепляет свободную ДНК между соседними нуклеосомами. Случайное расщепление хроматина ДНК-азой, сначала дает смесь частиц состоящих из одной или более нуклеосом содержащих приблизительно 200 пар оснований, каждая (рис. 2.4).
Как показали анализы состава индивидуальных нуклеопротеидных комплексов, мономерная нуклеосома содержит ДНК (~ 200 пар нуклеотидов) связанную с гистонами пяти типов. Участок ДНК, входящий в состав одной нуклеосомы, может несколько отличаться от усредненной величины в 200 пар нуклеотидов, которая была определена в первых экспериментах. Впоследствие оказалось, что в каждом конкретном случае получаемое значение достаточно точно соответствовало средней величине, которая чаще всего составляла 180-200 пар нуклеотидов. Однако были обнаружены и такие крайние значениия, как 260 пар нуклеотидов, например, в сперматозоидах морского ежа.
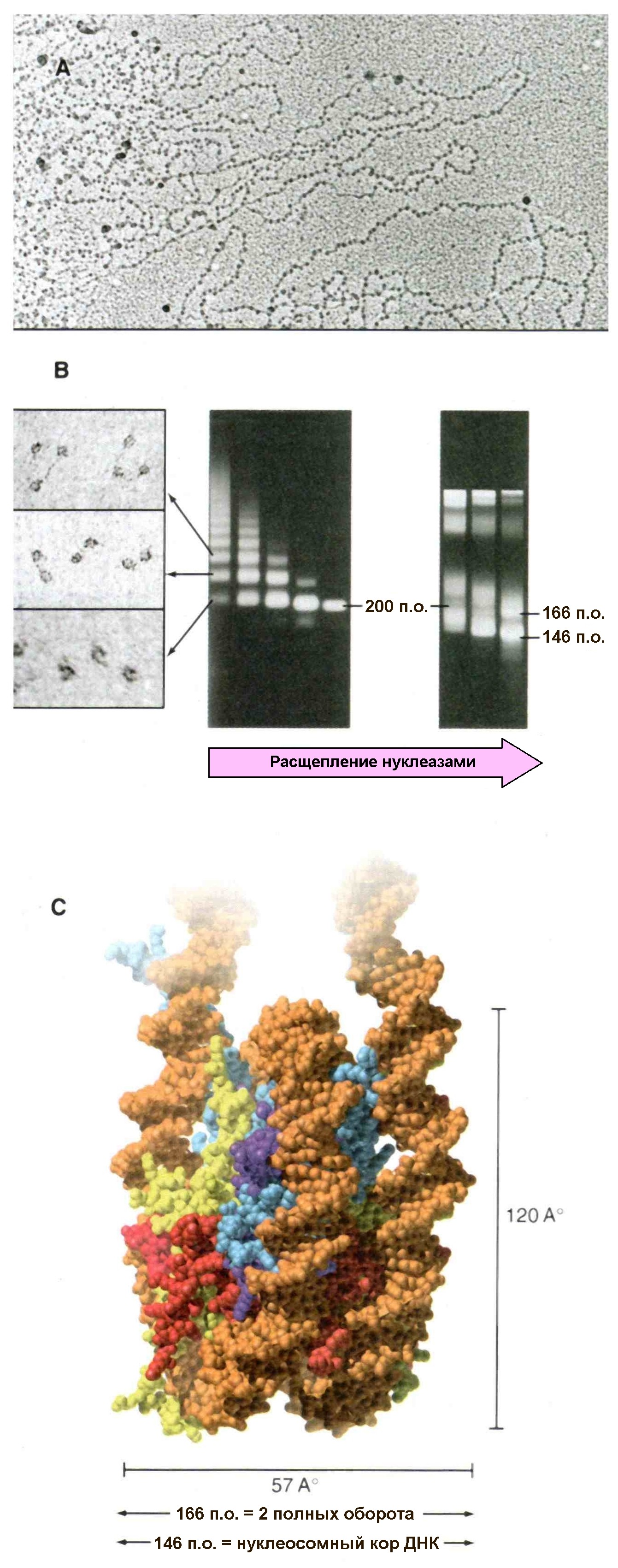
Рис. 2.4 |
Нуклеосомы. А – электронная микрофотография, показывающая петли хроматина покрытые нуклеосомами, которые при данных условиях выглядят как бусины на нити. В – под действием ДНК-азы из хроматина высвобождаются фрагменты, содержащие различное число нуклеосом (слева) причем длина ДНК входящей в состав нуклеосом составляет приблизительно 200 пар нуклеотидов (в центре). Более длительное воздействие ДНК-азы приводит к образованию минимальных нуклеосом (нуклеосомного кора), в которых участок ДНК представлен 146 парами нуклеотидов (справа). С – структура минимальной нуклеосомы по данным рентгеноструктурного анализа. ДНК накручена вокруг компактного гитонового кора (гистонового октамера). Woodcock C.L., Sweetman H.E., Frado L.L., Structural repeating units in chromatin. II Their isolation and partial characterization, Exp. Cell. Res., 97, 111-119, 1976, Allan J., Cowling G.J., Harborne N., et al., Regulation of the higher-order structure of chromatin by histones H1 and H5, J. Cell Biol., 90, 279-288, 1981, C – PDB file: 1 aoi. |
Поскольку все нуклеосомы состоят из одинакового набора гистонов, связанных с ДНК определенной длины, характерной для конкретного вида организма, возникает вопрос – существует ли в организации хроматина структура, отличающаяся неизменностью строения. Оказалось, что такая структура существует, и называется она минимальной нуклеосомой или нуклеосомным кором (core particle). Минимальная нуклеосома была идентифицирована по действию ДНК-азы микрококков на нуклеосомный мономер. Сначала фермент вносит разрезы в ДНК между нуклеосомами, а затем, если реакцию продолжить, нуклеаза подравнивает концы ДНК до образования минимальной нуклеосомы, при этом происходит ступенчатое уменьшение длины сегмента ДНК в составе данной структуры. Например, в ядрах клеток печени крыс нуклеосомные мономеры сначала содержали сегменты ДНК в 205 пар нуклеотидов. Дальнейшая обработка нуклеосом ДНК-азой приводит к образованию частиц включающих 166 пар оснований ДНК (такие частицы содержат два полных оборота ДНК вокруг белкового кора). Если продолжать обработку препарата ДНК-азой далее – образуются конечные стабильные частицы, содержащие 146 пар оснований, при этом ДНК делает 1,75 (1¾) оборота вокруг белкового кора (рис. 2.5).
Нуклеосомный кор, как было указано выше, состоит из сегмента хромосомной ДНК длиной 146 пар оснований намотанной на гистоновый октамер, включающий гистоны Н2А, Н2В, Н3 и Н4 – по две молекулы каждого белка. Изолированные минимальные нуклеосомы имеют дискообразную форму и представлены отрицательно суперспирализованной ДНК накрученной на октамер гистонового кора. Этот октамер состоит из центрального тетрамера построенного из двух прочно взаимодействующих гетеродимеров Н3:Н4. С каждой стороны тетрамера располагаются гетеродимеры Н2А:Н2В.
Гистоны составляют бóльшую часть оснóвных белков хроматина и присутствуют примерно в том же количестве (по массе), что и ДНК. По относительной доле оснóвных аминокислот, которую выражают отношением Lys/Arg, охарактеризовано пять типов гистонов, которые характеризуются разными размерами полипептидных цепей и разным аминокислотным составом (табл. 2.1).
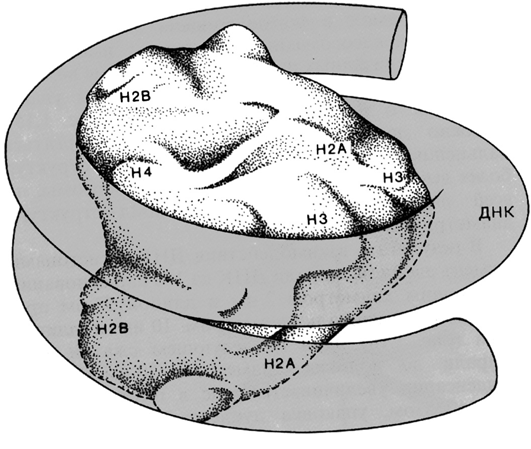
Рис. 2.5 |
Модель нуклеосомного кора, построенная по данным кристаллографического анализа низкого и высокого разрешения. Сегмент ДНК длиной 146 пар оснований обвивает гистоновый октамер, делая вокруг него 1¾ оборота (Kornberg R.D., Klug A., The nucleosome, Sci. American, 1981, 244, 48-62). |
Таблица 2.1