

Доказательства генетической роли нуклеиновых кислот
Методами формальной генетики было установлено, что ген является дискретным фактором наследственности, представляет собой часть хромосомы и, что он переходит от родителя к потомку, т.е. передается в ряду поколений.
Решающую роль в выяснении природы гена сыграло представление о том, что ген отличается от признака, который он определяет. До начала 40-х годов было лишь известно, что гены – это действительно основные единицы наследственности, но каким путем они выполняют свою функцию, оставалось неясным. Гены можно было характеризовать, исходя из мутаций, вызывающих некоторые отклонения от нормы в фенотипе. Почти во всех случаях эффект мутаций анализировали только на описательном уровне. Идея о том, что эффект мутаций связан с появлением измененного фермента возникла в начале 20-го столетия. В своих работах, проведенных в 1902-1908 годах, Арчибальд Гэррод высказал мнение, что болезнь человека – алкаптонурия – обусловлена нарушением какой-то метаболической реакции, катализируемой ферментом. Его книга «Врожденные ошибки метаболизма» была для своего времени самым ярким и важным вкладом в биологию и медицину. Фраза А. Гэррода – «расщепление бензольного кольца в ходе нормального метаболизма – результат действия специального фермента, а при врожденной алкаптонурии этот фермент отсутствует» – заключает в себе концепцию, согласно которой генетический дефект может привести к нарушению определенного метаболического процесса, что находит свое отражение в наблюдаемом фенотипе. Таким образом, к определенному моменту сформировались взгляды относительно того, что различные мутации влияют на фенотип посредством препятствия синтеза какого-либо белка. В 1941 году Дж. Бидл и Э. Тейтем, основываясь на данных по изучению мутантных штаммов хлебной плесени Neurospora crassa, сформулировали важнейший принцип – один ген – один белок (один фермент).
Физическая же и химическая природа генетического материала оставалась неизвестной.
Значительным толчком к изучению и пониманию природы гена послужил обзор Е. Шрëдингера, опубликованный в 1945 году. В обзоре Шрëдингером были развиты ранее высказанные соображения Макса Дельбрюка относительно того, что исходя из одних только законов физики, вряд ли можно объяснить свойства генетического материала, особенно его стабильность в ряду бесчисленных поколений. Никто не сомневался в том, что гены подчиняются законам физики, которые уже известны, но при этом думали, что изучение свойств генов может привести к открытию новых законов физики. Именно эта перспектива и привела многих физиков в биологию.
В обзоре Шрëдингер описал свойства генов таким образом, что с очень небольшими изменениями обрисованная им картина вполне соответствует современным представлениям: «Мы можем представить структуру гена в виде гигантской молекулы, способной только к локальному изменению, которое сводится к перестройке атомов и приводит к образованию изомерной молекулы. Перестройка [мутация] может повреждать только небольшую часть гена, но при этом возможно множество перестроек». И хотя Шрëдингер представлял себе ген как апериодический кристалл, поскольку тогда казалось, что это единственная физическая структура, способная удовлетворять требованиям, предъявляемым к генетическому материалу, именно эти ранние представления о гене как о гигантской молекуле объясняли, каким образом ген может быть протяженной частью хромосомы.
Несмотря на то, что ранние генетические исследования были сконцентрированы на высших организмах, первый случай переноса генетического материала обнаружили у бактерий. Хотя позднее тот же подход применили к высшим организмам и, в результате оказалось, что и у прокариот, и у эукариот генетический материал находится в одной и той же форме.
Самый первый и наиболее яркий пример, демонстрирующий роль ДНК, как вещества наследственности относится к 1928 году. Явление, получившее название трансформации впервые было обнаружено бактериологом Фредериком Гриффитом при изучении пневмококковой инфекции у мышей. Вирулентность пневмококков определяется капсульным полисахаридом, расположенным на поверхности клеточной стенки. Вирулентные бактерии образуют на поверхности агара гладкие колонии, обозначаемые буквой S (от англ. smooth – «гладкий»). Различные штаммы пневмококков обладают разными капсульными полисахаридами, но все они вирулентны. Любой штамм может дать мутантов, не способных к образованию соответствующих капсульных полисахаридов. Мутанты этих бактерий, утратившие способность к образованию полисахарида, невирулентны и образуют шероховатые колонии. Такие клетки обозначают буквой R (от англ. rough – «шероховатый»). Бескапсульные клетки легко фагоцитируются в организме мыши и потому не патогенны для животных. После термообработки S-клетки погибают и так же становятся невирулентными. Если же мышей инфицировать смесью живых R-клеток (невирулентных) и иниктивированных нагреванием S-клеток, то животные погибают от пневмококковой инфекции. В легких погибших животных были обнаружены вирулентные бактерии S. На основании полученных результатов Гриффит сделал вывод, что в убитых нагреванием S-клетках присутствует трансформирующий фактор, превращающий непатогенные живые R-клетки в вирулентные клетки S. Это изменение было стабильным: трансформированные пневмококки давали патогенное потомство S-формы. На рис. 3.1 приведена схема описанного выше эксперимента.
Впоследствии было установлено, что трансформация RS может успешно происходить в бесклеточной системе in vitro. Клетки растущей колонии R-формы трансформировались в S-форму при добавлении к ним бесклеточного экстракта из убитых нагреванием S-клеток (рис. 3.1). Это открытие положило начало исследованиям, направленным на установление химической природы трансформирующего фактора.
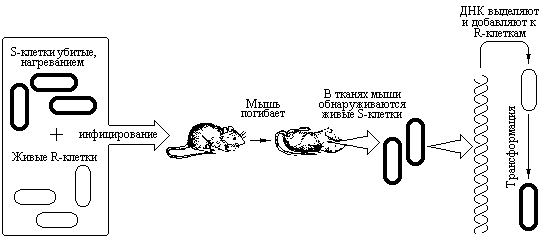
Рис. 3.1 Трансформация живых непатогенных бактерий R в вирулентные бактерии S. Инактивированные нагреванием S-клетки и живык R-клетки не патогенны для мыши. Однако, если мышь инфицировать смесью этих клеток, она погибнет и из легочной ткани могут быть выделены живые S-клетки. Трансформацию R-клеток в вирулентные S-клетки можно осуществить in vitro, если к R-клеткам добавить ДНК, выделенную из гладких бактерий (Lewin B., Genes III, New York, Wiley, 1987).
В 1944 году Освальд Эйвери, Колин Мак-Леод и Маклин Мак-Карти опубликовали результаты своих классических исследований [5]. Экспериментальное доказательство того, что трансформирующий фактор - это дезоксирибонуклеиновая кислота сводилось к следующим положениям:
элементный химический состав очищенного трансформирующего фактора был близок к рассчитанному элементному составу для ДНК;
оптические, электрофоретические свойства, поведение при ультрацентрифугировании соответствовали свойствам ДНК;
экстракция белков и липидов не вызывала потери трансформирующей активности;
обработка препарата трипсином и химотрипсином не влияла на трансформирующую активность;
обработка очищенного препарата РНК-азой не влияла на трансформирующее начало;
только обработка препарата ДНК-азой приводила к потере трансформирующей активности.
Таким образом, было показано, что в результате трансформации бактериальных клеток, именно ДНК определяет появление соединения отличного по химической природе от самой нуклеиновой кислоты (т.е. синтез капсульного полисахарида). Позднее были получены и другие результаты исследований по трансформации бактерий, указывающие на способность ДНК направлять синтез капсульных полисахаридов разных типов. Однако, даже после таких достаточно убедительных экспериментальных данных долгое время считали, что ДНК может определять биосинтез только полисахаридов и только у бактерий, но не является веществом наследственности вообще.
Следующее доказательство генетической роли ДНК было получено в 1952 году в экспериментах, проведенных на совершенно другой системе. Альфред Херши и Марта Чейз [6] инфицировали клетки E.coli фагом T2, у которого радиоактивным изотопом 32P была помечена ДНК, а изотопом 35S помечены белки капсида. Эксперимент был построен на том свойстве фага, что при добавлении его к бактериальной культуре он адсорбируется на поверхности клеток, вводит определенное вещество в бактериальную клетку и затем, через некоторое время клетка лизируется, давая большое число фаговых потомков.
Меченые фаги смешивали с клеточной суспензией E.coli и неадсорбированные фаговые частицы удаляли центрифугированием. Затем инфицированные бактериальные клетки подвергали сильному встряхиванию с помощью блендера и полученную суспензию разделяли на две фракции посредством центрифугирования. Супернатантная фракция содержала пустые фаговые оболочки, отделившиеся от клеточных стенок бактерий, а осадок состоял из самих инфицированных бактериальных клеток. В результате анализа распределения изотопов по фракциям оказалось, что практически вся метка 35S сосредоточена в супернатанте, в то время как большая часть метки 32P оказалась внутри бактериальных клеток. После лизиса клеток E.coli в потомстве фага Т2 было обнаружено 30% исходной метки 32P и менее 1% метки 35S. Этот эксперимент прямо показывает, что меченая родительская фаговая ДНК проникает в бактерию и затем, обеспечивая размножение фага, становится частью фагового потомства (см. рис. 3.2).
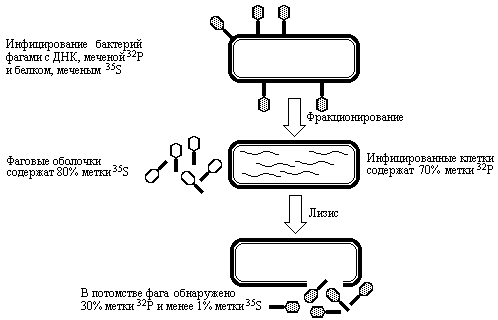
Рис. 3.2 Генетическим материалом фага Т2 является ДНК. ДНК фаговых частиц, меченая 32Р, после включения в бактериальные клетки обнаруживается в фаговом потомстве, тогда как метка 35S у потомков фага не идентифицируется (Lewin B., Genes III, New York, Wiley, 1987).
Необходимо обратить внимание на следующее обстоятельство. Хотя эксперименты А. Херши и М. Чейз доказательны в отношении генетической роли ДНК, в них не так полно, как в опытах по трансформации, исключается возможность загрязнения используемого препарата белковыми примесями. В опытах по трансформации, используя совершенные методы очистки ДНК, можно свести содержание белковых примесей до ничтожно малой величины. При инфицировании фагом в бактериальную клетку вместе с ДНК неизбежно попадает некоторое небольшое количество белка. Однако, поскольку со времени работ О. Эйвери точка зрения биологов в отношении ДНК, как вещества наследственности изменилась, результаты экспериментов А. Херши и М. Чейз были сразу восприняты как доказательство генетической роли ДНК.