

Практическое занятие № 1, 2. Самореплицирующиеся молекулы. Естественный отбор самореплицирующихся молекул.
Любой процесс копирования неизбежно связан с возникновением ошибок, приводящих к размножению неточных копий оригинала. Следовательно, при многократных циклах репликации полинуклеотида, образующаяся последовательность будет существенно отличаться от исходной. Таким способом формируется огромное разнообразие молекул. В случае РНК эти молекулы будут характеризоваться и различными функциональными свойствами, поскольку последовательность оснований - не просто цепочка символов. Молекулы РНК, обладающие разной нуклеотидной последовательностью, должны обладать выраженной химической индивидуальностью, влияющей на их поведение. Каждая измененная молекула в соответствии с конкретной последовательностью будет приобретать в растворе вполне определенную конформацию. Дело в том, что основания в составе РНК могут не только спариваться со свободными нуклеотидами среды, но могут образовывать комплементарные пары в пределах самой молекулы РНК.
Из-за подобных взаимодействий возникают различные трехмерные структуры, и каждая молекула в целом приобретает уникальную конформацию, полностью зависящую от специфической нуклеотидной последовательности. Схематическое изображение одного из таких теоретически возможных конформеров приведено на рис. 1.4. В свою очередь, пространственная трехмерная укладка полинуклеотида не может не влиять на его стабильность и на способность реплицироваться. Поэтому далеко не все молекулы в реплицирующейся смеси одинаково успешно смогут размножаться. В лабораторных экспериментах было показано, что реплицирующиеся молекулы РНК подвержены своеобразному естественному отбору, в результате которого в зависимости от создаваемых условий начинает преобладать та или иная последовательность.
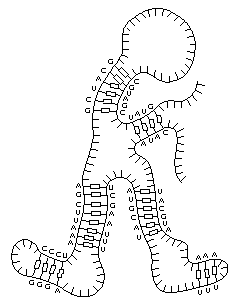
Рис. 1.4 Возможная трехмерная структура молекулы РНК. В результате спаривания нуклеотидов, расположенных в разных участках одной и той же рибополинуклеотидной цепи, вся молекула приобретает определенную конформацию (Alberts B. et al., Molecular biology of the cell, New York, Garland, 1989).
Таким образом, молекула РНК обладает двумя важнейшими свойствами: - закодированная в последовательности нуклеотидов информация передается в процессе репликации, а уникальная пространственная структура определяет характер взаимодействия с другими полинуклеотидными молекулами и реакцию на внешние условия. Оба этих свойства - информационное и функциональное - являются необходимыми предпосылками эволюционного процесса.
Для каждой реплицирующейся молекулы РНК критическим компонентом среды является набор других молекул РНК в системе. Кроме того, что эти молекулы являются матрицами, определяющими собственную репликацию, они могут катализировать следующие процессы:
образование и разрушение ковалентных связей, в том числе и между нуклеотидами;
некоторые специализированные молекулы РНК могут катализировать изменения в других молекулах РНК, разрезая нуклеотидную последовательность в определенной точке (РНК-компонент рибонуклеазы Р из E. coli способен самостоятельно проявлять каталитическую активность в процессе созревания тРНК);
другие типы РНК могут вырезать часть своей собственной последовательности и сшивать разрезанные концы (самосплайсинг рРНК у Tetrahymena thermophila).
Можно предположить, что некоторые из перечисленных реакций имели первостепенное значение в первичном бульоне. Например, определенная каталитическая РНК, действуя на собственные копии, обеспечивала свою репликацию более эффективно. С другой стороны, находящиеся в растворе другие молекулы РНК, могли способствовать репликации первой молекулы. Если эти благоприятные эффекты взаимосвязаны, то различные типы РНК, обладающие той или иной специфичностью в отношении катализируемой реакции, должны были сформировать кооперативную систему, которая реплицировалась с необычайно высокой эффективностью. Схема трех последовательных стадий эволюции самореплицирующейся системы молекул РНК, способных направлять даже синтез белков приведена на рис. 1.5.
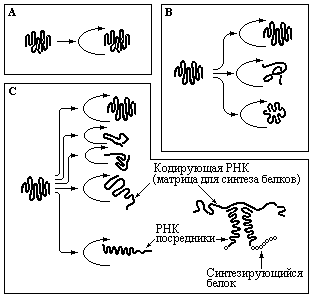
Рис. 1.5 А – каталитическая молекула РНК соединяет нуклеотиды для воспроизведения собственной нуклеотидной последовательности. В – семейство каталитических молекул РНК, в котором одна из них преимущественно катализирует репликацию других РНК. С – появление новых каталитических РНК, способных присоединять к себе активированные аминокислоты. Вследствие комплементарного спаривания с кодирующей молекулой РНК последовательность таких РНК действует как матрица для синтеза полимеров из аминокислот, в результате чего появляются первые генетически детерминированные последовательности белков. Таким образом, РНК служат первыми посредниками между последовательностями нуклеотидов и аминокислот (Alberts B. et al., Molecular biology of the cell, New York, Garland, 1989).
Итак, предполагается, что 3,5-4 млрд. лет назад самореплицирующиеся системы молекул РНК положили начало эволюционному развитию на Земле. Системы молекул РНК с различными наборами последовательностей конкурировали за запасы предшественников, необходимых им для построения копий (аналогично конкуренции организмов за пищевые ресурсы в современных условиях). Успех «выживаемости» каждой самореплицирующейся системы зависел от точности и скорости копирования, а также от стабильности копий.
Несмотря на то, что структура полинуклеотидов хорошо приспособлена для хранения и передачи информации, каталитические возможности РНК весьма ограничены. Значительно бóльшая универсальность присуща полипептидам, состоящим из 20 аминокислот, которые обладают химически разнообразными боковыми радикалами и способны принципиально принимать неисчислимое множество различных пространственных форм, обладающих многообразными реакционноспособными участками.
Можно считать весьма вероятным, что полипептиды со случайными аминокислотными последовательностями, возникавшие под воздействием пребиотических синтетических механизмов, уже могли проявлять каталитические свойства и, в частности, могли облегчать репликацию РНК. С другой стороны, те полинуклеотиды, которые способствовали синтезу полезных для них полипептидов должны были приобрести большее преимущество. В главе «Трансляция» будет рассмотрен биохимический аппарат современной клетки, осуществляющий синтез белка. В настоящее время механизм синтеза полипептидных цепей характеризуется высокой степенью сложности, а сам процесс нуждается в участии различных макромолекул (иРНК, рРНК, тРНК, ферментативный аппарат) и белковых факторов.
На сегодняшний день представляется вероятным, что РНК примитивным способом могла обеспечивать первичный синтез белков. Очевидно в пребиотических условиях, предшественники информационных и транспортных РНК направляли примитивный синтез полипептидов без помощи белков (рис. 1.5 С). Каталитическая функция молекул РНК подтверждается тем фактом, что в современных рибосомах рибосомной РНК принадлежит роль главного катализатора процесса синтеза белка.
Синтез специфических белков под управлением РНК потребовал «разработки» кода, определяющего соответствие (коллинеарность) последовательности нуклеотидов последовательности аминокислот в белке. Код был «выбран», вероятно, случайно и до сих пор остается фактически одинаковым у всех живых организмов. Это наводит на мысль, что все современные клетки являются потомками одной примитивной линии клеток, сумевших разработать эффективный механизм синтеза необходимых ей белковых катализаторов.
Как только эволюция нуклеиновых кислот продвинулась до кодирования ферментов, обеспечивающих их собственное воспроизведение, распространение репликативной системы должно было резко ускориться.
Одним из решающих событий, приведших к формированию первой клетки, очевидно, было формирование внешней мембраны. Вполне понятно, что белки, синтезируемые под контролем РНК определенного типа не могли бы в полной мере облегчить воспроизведение именно этих молекул РНК, если бы не находились поблизости от них. Более того, до тех пор, пока белки свободно диффундировали среди реплицирующихся молекул РНК, они, в принципе, могли способствовать размножению любого типа конкурирующей РНК. Если возникала РНК, кодирующая лучший тип синтезирующего фермента, то он был лишен возможности именно избирательно обеспечивать выживание этой измененной РНК. Отбор молекул РНК по качеству создаваемых ими белков не мог начаться раньше, чем появился некий замкнутый компартмент, заключающий в себе и кодирующую РНК и соответствующие белковые продукты.
Важнейшая роль в эволюции клеточных мембран принадлежит классу амфипатических соединений, обладающих простым физико-химическим свойством: одна часть молекулы гидрофобна, а другая – гидрофильна. Когда такие соединения находятся в водной фазе, они спонтанно агрегируют с образованием бислойных структур в виде небольших везикул, в которых гидрофобные части молекул приходят в контакт друг с другом, а гидрофильные части контактируют с водой. Таким образом бислойные везикулы изолируют водное содержимое от внешней среды. Если эффективно реплицирующаяся РНК заключена в липидном компартменте, то любой кодируемый ею белок будет использоваться для повышения скорости и точности копирования только этой РНК.
Все ныне существующие клетки окружены плазматической мембраной, состоящей из амфипатических молекул, представленных главным образом фосфолипидами.
Не совсем ясно в какой момент эволюции биологического катализа были сформированы клетки. Они могли появиться, когда молекулы фосфолипидов пребиотического бульона случайно собирались в мембраноподобные структуры, которые также случайно заключали в себя самореплицирующиеся смеси каталитических молекул РНК. Однако, принято считать, что катализируемый примитивный синтез белков осуществлялся до появления клеток. В любом случае, как только самореплицирующиеся смеси каталитических молекул РНК и соответствующих белков оказались заключенными в замкнутую мембрану, молекулы РНК начали эволюционировать не только на основе совершенствования своей структуры и функции, но также воздействуя на другие молекулы компартмента. Нуклеотидные последовательности РНК могли теперь влиять на признаки всего компартмента, т.е. целой клетки.
Первые клетки на Земле, по-видимому, содержали очень небольшое количество компонентов и делились значительно медленнее современных. Существует и более значительное отличие между примитивными клетками и ныне существующими: – генетическая информация в современных клетках хранится в ДНК, а не в РНК. В современных клетках присутствуют оба типа нуклеиновых кислот, но в ходе эволюции они специализировались, выполняя свои уникальные функции, каждая. Незначительные химические отличия делают их приспособленными для решения разных задач.
Дезоксирибонуклеиновая кислота используется для хранения генетической информации по ряду причин:
- молекула ДНК химически более стабильна, чем молекула РНК;
- в молекуле ДНК отсутствует одна гидроксильная группа, что делает ее более устойчивой к гидролизу, чем РНК;
- ДНК существует в виде двухцепочечной молекулы, состоящей из двух комплементарных полинуклеотидных цепей, что позволяет ДНК легко реплицироваться и репарировать возникающие в ней повреждения, при этом неповрежденная цепь используется в качестве матрицы для восстановления комплементарной дефектной цепи;
- используя принцип комплементарности, ДНК направляет синтез отдельных видов молекул РНК. Синтезированные таким способом одноцепочечные молекулы РНК выполняют две другие функции первобытных полинуклеотидов: одни направляют синтез белков, как кодирующие молекулы (информационные РНК) , а другие, как каталитические молекулы (рибосомные и другие неинформационные РНК).
Существующие на сегодняшний день представления об эволюции первобытных молекул можно выразить следующим образом. Генетические и каталитические свойства РНК позволяют предположить, что именно эти молекулы первыми включились в эволюцию. После возникновения достаточно эффективного механизма синтеза белка ДНК приняла на себя генетическую функцию, при этом белки стали катализаторами, а РНК сохранилась, главным образом, как промежуточное звено между ними. ДНК стала необходимой в качестве главной информационной молекулы только тогда, когда клетки значительно усложнились и для них потребовалось больше генетической информации, чем та, которую могли стабильно обеспечить молекулы РНК.
(2) Нуклеиновые кислоты. Строение азотистых оснований. Пуриновые и пиримидиновые основания. Строение нуклеозидов и нуклеотидов.
Нуклеиновые кислоты выполняют ряд важных биологических функций, не свойственных другим полимерным веществам. В частности, они обеспечивают хранение и передачу наследственной информации и принимают непосредственное участие в механизмах реализации этой информации путем программирования синтеза всех клеточных белков. Структурные компоненты нуклеиновых кислот выполняют, кроме того, функции кофакторов (коэнзим А, уридин-дифосфатглюкоза и др.), аллостерических эффекторов, входят в состав коферментов (никотинамидадениндинуклеотид, флавинадениндинуклеотид и др.), принимая тем самым непосредственное участие в обмене веществ, а также в аккумулировании (накоплении), переносе и трасформации энергии. Они являются предшественниками вторичных посредников (мессенджеров) – циклических мононуклеотидов (цАМФ и цГМФ), выполняющих важную функцию в передаче внутриклеточных сигналов.
О локализации и количественном содержании нуклеиновых кислот
в клетках получены определенные данные. Доказано, что количественное
содержание ДНК в клетках одного и того же организма отличается
удивительным постоянством и исчисляется несколькими пикограммами,
однако в клетках разных видов живых организмов имеются существенные
количественные различия в содержании ДНК. Хорошо известно также, что
ДНК преимущественно сосредоточена в ядре, а в митохондриях и хлоро-
пластах содержится только небольшой процент клеточной ДНК. О количестве РНК нет точных данных, поскольку содержание ее в разных клетках в значительной степени определяется интенсивностью синтеза белка. На долю РНК приходится около 5–10% от общей массы клетки. Современная классификация различных типов клеточной РНК основывается на данных топографии, функции и молекулярной массы. Выделяют три главных вида РНК: матричную (информационную) – мРНК, которая составляет 2–3% от
всей клеточной РНК; рибосомную – рРНК, составляющую 80–85% и транспортную – тРНК, которой около 16%. Эти 3 вида различаются нуклеотидным составом и функциями.
Матричная РНК (мРНК) синтезируется в ядре на матрице ДНК, затем поступает в рибосому, выполняя матричную функцию при синтезе белка.
Транспортные РНК (тРНК) имеют небольшую молекулярную массу и содержатся в растворимой фракции цитоплазмы, выполняя функцию переноса аминокислот к месту белкового синтеза – рибосоме. Рибосомные РНК (рРНК) имеют разную и значительно большую молекулярную массу. Они локализуются в двух субчастицах рибосом 50S и 30S у Е.coli и 60S и 40S в клетках животных.
Субчастица 60S содержит три разных рРНК (5S, 5,8S и 28S рРНК), в то время как субчастица 40S – одну молекулу 18S рPHK. (Размер субчастицы измеряется в ед. Сведберга (S) - 60 (более 50 белков) и 40 (более 30 белков)
Нуклеиновые кислоты (ДНК и РНК) относятся к сложным высокомолекулярным соединениям, это линейные нерегулярные полимеры, мономерами которых служат нуклеотиды. Каждый нуклеотид (рибонуклеотид или дезоксирибонуклеотид) состоит из трех частей: 1) остатка фосфорной кислоты, 2) пятиуглеродного моносахарида в циклической форме, 3) азотистого основания.
В молекуле ДНК углевод представлен дезоксирибозой, а в молекуле РНК – рибозой, отсюда их названия: дезоксирибонуклеиновая (ДНК) и рибонуклеиновая (РНК) кислоты. Кроме того, они содержат фосфорную кислоту, по два пуриновых и по два пиримидиновых основания; различия только в пиримидиновых основаниях: в ДНК содержится тимин, а в РНК –
урацил.
Углеводы (рибоза и дезоксирибоза) в молекулах ДНК и РНК находятся
в β-D-рибофуранозной форме:
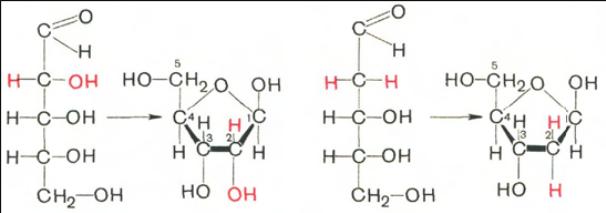
D-рибоза D-рибофураноза D-2-дезоксирибоза D-2-дезоксирибофураноза
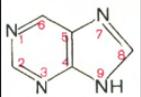
Основу структуры пуриновых и пиримидиновых оснований составляют
два ароматических гетероциклических соединения – пиримидин и пурин:
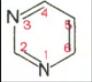
Пиримидин Пурин
Молекула пурина состоит из двух конденсированных колец: пиримидина и имидазола.
В составе нуклеиновых кислот встречаются три главных пиримидиновых основания: цитозин, урацил и тимин.
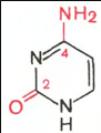
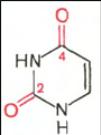
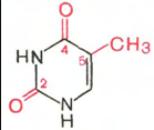
Цитозин Урацил Тимин
Два пуриновых основания, постоянно встречающихся в гидролизатах нуклеиновых кислот, имеют следующее строение:
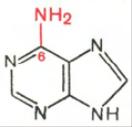
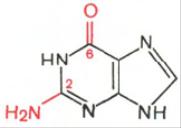
Аденин Гуанин
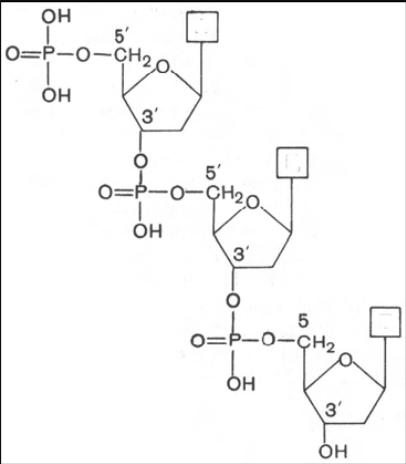
Первичная структура ДНК. Последовательно расположенные нуклеотиды в молекулах ДНК и РНК ковалентно связаны между собой в цепь через остаток фосфорной кислоты: 5'-гидроксильная группа моносахарида одного нуклеотида присоединена к 3'-гидроксильной группе моносахарида соседнего нуклеотида с помощью фосфодиэфирной связи..
Пуриновое или пиримидиновое основание в нуклеотидах соединяется с углеводом N-гликозидной связью. В составе нуклеиновых кислот обнаруживаются только β-нуклеотиды. Примером могут служить два мононуклеотида: аденозин-5'-монофосфорная кислота (АМФ) и цитидин-5'-монофосфорная кислота (ЦМФ):
R у 2' углерода представлен Н- или ОН-группой в зависимости от типа нуклеиновой кислоты – ДНК или РНК. В образовании N-гликозидной связи в пуриновых нуклеотидах принимают участие N-9 пурина и С-1' пен-
тозы,а в пиримидиновых нуклеотидах – N-1 пиримидина и С-1' пентозы.
Чтобы отличить углеродные атомы рибозы или дезоксирибозы от углеродных атомов, входящих в состав пуриновых и пиримидиновых оснований, первые принято обозначать символом ≪штрих≫: например, атомы у 3-го и 5-го углерода обозначают С-3' и С-5' или, чаще, 3' и 5'.
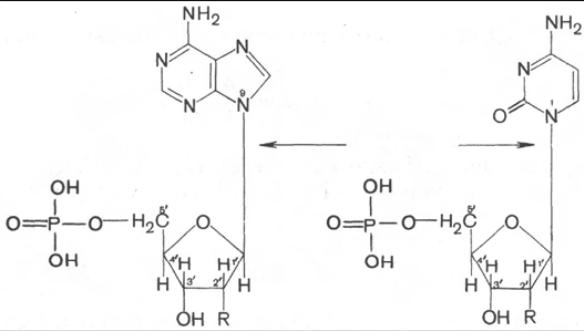
АМФ ЦМФ
Таким образом, ковалентные остовы нуклеиновых кислот состоят из монотонно чередующихся фосфатных и сахаридных групп; основания
же можно рассматривать как боковые группы, присоединенные к остову
на равных расстояниях друг от друга. Цепи ДНК и РНК обладают определенной полярностью, или направлением, поскольку все межнуклеотидные фосфодиэфирные связи ориентированы вдоль цепи одинаково. Благодаря этой полярности каждая полинуклеотидная цепь имеет 5'-конец и 3'-конец. Сахарофосфатный остов и ДНК, и РНК несет отрицательный заряд, поскольку фосфатные группы депротонированы при
характерных для клеток pН.
Нуклеотиды – фосфорные эфиры нуклеозидов, которые построены из остатка углевода – пентозы и азотистого основания.
Состав нуклеозидов и мононуклеотидов
Азотистые основания
|
Нуклеозиды (основание + углевод)
|
Мононуклеотиды (нуклеозиды + фосфорная кислота)
|
Сокращенное обозначение
|
Аденин |
Аденозин |
Аденозинмонофосфат (адениловая кислота)
|
АМФ |
Гуанин |
Гуанозин |
Гуанозинмонофосфат (гуаниловая кислота)
|
ГМФ |
цитозин |
Цитидин |
Цитидинмонофосфат (цитидиловая кислота)
|
ЦМФ |
урацил |
Уридин |
Уридинмонофосфат (уридиловая кислота)
|
УМФ |
тимин |
тимидин |
Тимидинмонофосфат (тимидиловая кислота)
|
ТМФ |
Вторичная структура ДНК.
В соответствии с моделью Дж. Уотсона и Ф. Крика, предложенной в 1953 г. на основании ряда аналитических данных, а также рентгеноструктурного анализа молекула ДНК состоит из двух цепей, образуя правовращающую спираль, в которую обе полинуклеотидные цепи закручены вокруг одной и той же оси. Удерживаются цепи благодаря водородным связям, образующимся между их азотистыми основаниями. Обе цепи полинуклеотидов в биспиральной молекуле ДНК имеют строго определенное пространственное расположение, при котором азотистые основания находятся внутри, а фосфорильные и углеводные компоненты – снаружи. Азотистые основания образуют между собой водородные связи по принципу комплементарности: между аденином и тимином образуется две водородные связи, между гуанином и цитозином -3. Реакции взаимодействия гуанина с цитозином и аденина с тимином получили название спаривания оснований. Количественное соотношение пуриновых и пиримидиновых азотистых оснований подчиняется правилам Чаргаффа:
Состав ДНК различных клеток, составляющих ткани и органы одного организма, всегда одинаков;
состав ДНК клеток организма с возрастом не изменяется;
состав ДНК клеток разных видов различен;
количество аденина всегда равно количеству тимина, а количество гуанина равно количеству цитозина;
сумма пуриновых оснований равна сумме пиримидиновых оснований (А+Г=Ц+Т или А+Ц=Г+Т).
Согласно модели Уотсона и Крика две цепи в молекуле ДНК –антипараллельны, т.е. идут в противоположных напавлениях. Поэтому, рассматривая спираль вдоль оси, можно видеть, что одна цепь идёт в направлении 5'–>3', а другая– 3'–>5'. Основания имеют плоскую форму и располагаются парами в плоскости, перпендикулярной оси спирали. Вдоль спирали основания уложены стопками друг на друга. Спиральная структура ДНК стабилизируется благодаря стэкинг-взаимодействиям, возникющим между ароматическими кольцами азотистых оснований.
Каждая пара оснований повёрнута на 36 градусов вокруг оси спирали относительно следующей пары, и таким образом 10 пар оснований составляют полный оборот в 360 градусов. Две цепи, образующие двойную спираль, уложены такимспособом, что наблюдаема структура характеризуется наличием малой бороздки шириной 12 А и большой бороздки шириной 22 А. Двойная спираль правозакрученная: если смотреть вдоль оси спирали – повороты следуют по часовой стрелке. Данное описание соответствует модели ДНК, известной как В-форма. Методами рентгеноструктурного анализа доказано существование других форм ДНК, названных А-, С-, D-, Е- и Z-формами (альтернативные структуры ДНК). У А-формы наблюдается некоторое смещение пар оснований от оси молекулы к периферии, что отражается на размерах (2,8 нм – длина одного витка, в котором вместо 10 содержится 11 мононуклеотидов; меняется расстояние между нуклеотидами и др.). Если А- и В-формы представляют собой правозакрученную двойную спираль, то Z-форма (зигзагообразная) ДНК имеет левозакрученную конфигурацию, в которой фосфодиэфирный остов располагается зигзагообразно вдоль оси молекулы. В настоящее время есть основание считать, что между А- и В-формами ДНК осуществляются взаимные переходы при изменении концентрации соли и степени гидратации. В этих переходах, которые могут быть вызваны растворителями или белками, очевидно, заключен определенный биологический смысл. Предполагают, что в А-форме ДНК выполняет роль матрицы в процессе транскрипции (синтез РНК на молекуле ДНК), а в В-форме – роль матрицы в процессе репликации (синтез ДНК на молекуле ДНК).