

Явление суперспирализации днк.
Двухцепочечные кольцевые молекулы ДНК, за крайне редким исключением, обладают уникальными топологическими характеристиками. Кольцевые молекулы имеют в структуре соответствующие изгибы и петли, которые получили название супервитков и которые хорошо различимы при использовании электронной микроскопии (рис. 3.13).
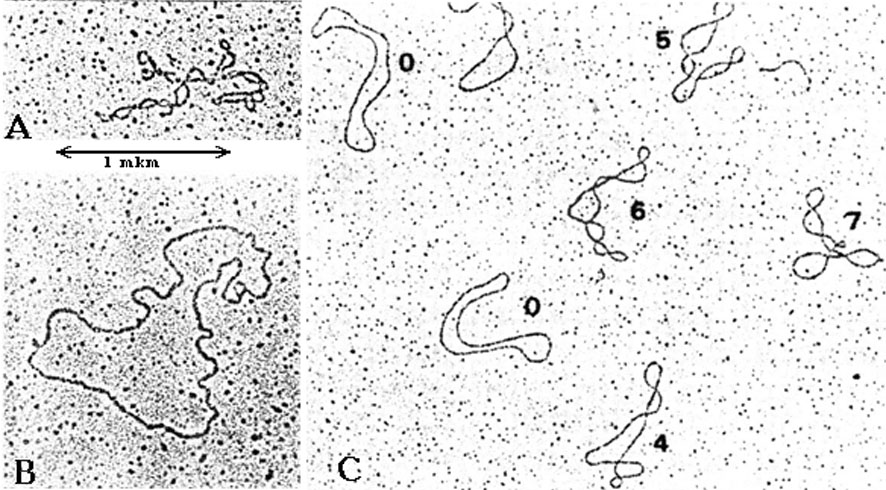
Рис. 3.13 |
Электронные микрофотографии суперспирализованных молекул ДНК: А – суперспирализованная митохондриальная ДНК; В - релаксированная форма митохондриальной ДНК (Страйер Л., Биохимия, т. 3, Мир, М., 1985, с. 21); С - двухцепочечная кольцевая ДНК фага М13 с разной степенью сверхспиральности. На последней микрофотографии цифрами указано число супервитков в каждой молекуле. (Сингер М., Берг П., Гены и геномы, т. 2, Мир, М., 1998, с. 46). |
Для того, чтобы максимально ясно представить происхождение этих супервитков может оказаться полезным рассмотрение двух моделей, в соответствии с которыми любая линейная молекула ДНК способна приобретать кольцевую форму. Кольцевая структура ДНК может сформироваться за счет сближения и ковалентного соединения соответствующих 3/- и 5/-концевых групп линейной молекулы с образованием фосфодиэфирных связей. Если при этом с данной молекулой ДНК не производить никаких манипуляций, результатом соединения концов будет образование релаксированной кольцевой ДНК, т.е. такой кольцевой двухцепочечной структуры, которая находится в термодинамически выгодном состоянии и характеризуется параметрами двойной спирали в классической В-форме, а это означает, что такая спираль включает 10 пар нуклеотидов на один полный оборот. Однако, если перед соединением концов линейной ДНК один из «хвостов» молекулы зафиксировать, а другой повернуть на один или несколько полных оборотов в направлении раскручивания двойной спирали, то в результате получим напряженную структуру. Такая напряженная структура, которая характеризуется недостатком оборотов получила название отрицательно суперспиролизованной ДНК. Возникшие отрицательные супервитки закручивают ДНК против часовой стрелки, т.е. в направлении, противоположном ходу обычной правосторонней двойной спирали. Такую спираль называют недокрученной. Вполне понятно, что данное явление не может происходить спонтанно, а должно инициироваться под действием внешних сил. Этими внешними силами являются специфические ферменты геликазы и топоизомеразы. Следует иметь в виду, что отрицательная суперспирализация снижает напряжение скручивания двойной спирали и, в ряде случаев, может приводить к разделению цепей.
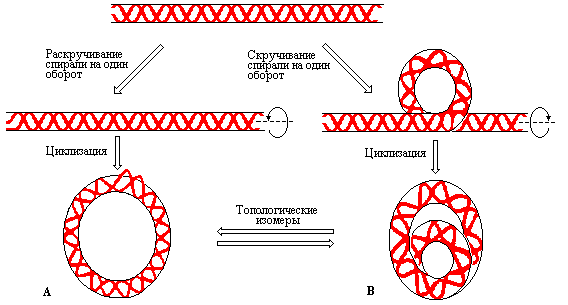
Рис. 3.14 |
Образование напряженных структур с отрицательной (А) и положительной (В) суперспирализацией (по рисунку из Textbook of Biochemistry with clinical correlations, Devlin T.M., ed., WILEY-LISS, 1993). |
Торсионное напряжение, неизбежно возникающее в таких суперспирализованных молекулах приводит к увеличению стандартной свободной энергии ДНК приблизительно на 10 ккалмоль на каждый супервиток, вводимый в структуру. Следовательно, супервитки можно рассматривать как форму запасания энергии. Практически это означает, что отрицательная суперспирализация должна влиять на стабильность структуры. Действительно, суперспирализованная ДНК подвергается структурным изменениям в таких условиях, при которых релаксированная ДНК остается без изменений. Напряжение, возникающее вследствие дефицита витков, может быть компенсировано, как было упомянуто выше, разрушением водородных связей между комплементарными парами оснований и раскрытием двойной спирали на небольшом участке макромолекулы. В результате разрушения водородных связей получаемая структура может приобрести вид регулярной релаксированной двойной спирали, содержащей небольшую одноцепочечную петлю называемую «пузырьком». Отсюда следует, что излишек энергии, которым обладают отрицательно суперспирализованные молекулы, может быть использован для разделения цепей ДНК. С другой стороны, водородные связи между парами оснований не обязательно должны разрываться. В этом случае, чтобы также смягчить напряжение, введенное в молекулу за счет раскручивания, кольцевая ДНК будет сворачиваться в направлении, противоположном тому, в котором она была повернута до циклизации. Процесс образования отрицательно суперспирализованной ДНК показан на рис. 3.14.
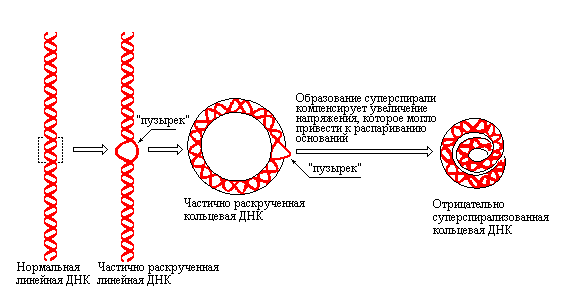
Рис. 3.15 |
Образование супервитков компенсирует возможное локальное разделение цепей ДНК, вызванное торсионным напряжением раскручивания (по рисунку из Textbook of Biochemistry with clinical correlations, Devlin T.M., ed., WILEY-LISS, 1993). |
Таким образом, напряжение вращения, которое было внесено в молекулу перед формированием кольцевой структуры может быть смягчено за счет образования третичной структуры с хорошо различимыми супервитками. Описанные два варианта последствий отрицательной суперспирализации не должны рассматриваться, как два совершенно различных типа суперспиралей, а представляют собой два проявления одного и того же феномена. В целом, то или иное состояние молекулы ДНК определяется ее окружением и соотношением степени нарушения водородных связей между парами оснований, с одной стороны, и степенью сверхскрученности, с другой стороны. На практике это означает, что отрицательно суперскрученная ДНК состоит из свернутых структур с повышенной склонностью к образованию неспаренных участков.
В любой кольцевой ДНК, которая исходно находится в релаксированной форме существует тенденция к образованию супервитков. Если же ДНК суперспирализована, то эта молекула будет тем сильнее стремиться к снижению напряженности, чем выше в ней плотность супервитков. Все существующие в природе молекулы ДНК имеют дефицит спирализованности, т.е. существуют в виде отрицательных суперспиралей, характеризующихся приблизительно одинаковой плотностью витков для всех типов ДНК независимо от источника. Как правило, все молекулы ДНК имеют, в среднем, один отрицательный виток на каждые 20 оборотов двойной спирали.
Вращение же линейного дуплекса в обратном направлении дает положительную суперспираль. Если до превращения линейной ДНК в соответствующую кольцевую стркутуру один из концов линейного двухцепочечного полинуклеотида поворачивать в направлении скручивания, а не раскручивания двойной спирали, то в результате этого получим кольцевую ДНК, содержащую положительные супервитки. Такую ДНК называют перекрученной (рис. 3.14). Положительно суперспирализованная ДНК не может снизить напряжение в структуре. В то время как отрицательно сверхспирализованные ДНК могут обеспечить снятие напряжения либо посредством разрушения водородных связей, либо за счет образования скрученных структур, единственным доступным способом снятия торсионного напряжения в положительно суперспирализованной ДНК является приобретение соответствующей третичной структуры (рис. 3.4 В). Положительную сверхспирализацию в молекуле ДНК можно получить экспериментально различными воздействиями in vitro, например, с помощью ферментов ДНК-топоизомераз, однако в естественных условиях такой способ сверхспирализации в ДНК встречается крайне редко и является преходящим.
Представления о суперспирализации часто бывают сложными для понимания без рассмотрения подходящей физической модели. В отсутствие более удачного примера можно попытаться представить происходящие события скрутив два отрезка тонкой натянутой веревки. В данном случае скрученная веревка будет представлять собой положительную суперспираль, которая для снятия напряжения от сверхспирализации, после того как сильное натяжение будет устранено, приобретет форму компактного завитка. Если при этом отпустить один из концов двух отрезков веревки, система разрушится и суперспираль приобретет релаксированную форму.
Как уже упоминалось, кольцевые отрицательно суперспирализованные молекулы ДНК могут приобретать несколько конформаций находящихся в динамическом равновесии друг с другом. Эти структуры характеризуются практически одинаковыми «параметрами напряжения». На рис. 3.15 приведены две альтернативные структуры отрицательно суперспирализованной кольцевой ДНК.
Вместе с тем, было показано, что помимо кольцевой структуры с «пузырьком» и тороидальной формы кольцевая отрицательно суперспирализованная ДНК может существовать также в виде третьей эквивалентной структуры, называемой «перекрещенной» и имеющей форму восьмерки (рис. 3.16).
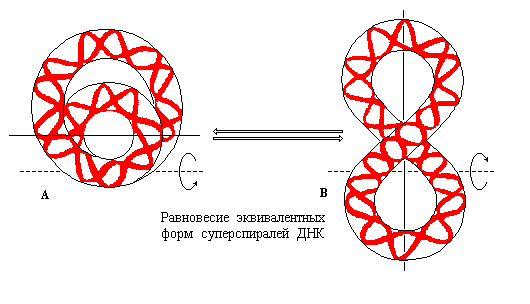
Рис. 3.16 |
Альтернативные равновесные структуры отрицательно суперспирализованной ДНК. А – тороидальная форма, В – «перекрещенная» структура (по рисунку из Textbook of Biochemistry with clinical correlations, Devlin T.M., ed., WILEY-LISS, 1993). |
Хорошо известно, что у мелких вирусов, большей части бактериофагов, вирусов животных, в клетках прокариот (хромосомы и плазмиды) и в клеточных органеллах эукариот ДНК находится в виде кольцевых молекул, содержащих отрицательные супервитки. Несмотря на то, что кольцевые молекулы ДНК, в первую очередь, являются идеальными претендентами на приобретение отрицательно суперспирализованной структуры in vivo, любой фрагмент двухцепочечной ДНК, у которой один из концов иммобилизован, в принципе, способен к суперспирализации. Поэтому можно справедливо полагать, что это свойство не является исключительной прерогативой только кольцевых ДНК. Любая соответствующим образом «заякоренная» линейная ДНК может приобрести суперспирализованную конформацию. Например, ДНК животных клеток в норме связанная с ядерными белками попадает в эту категорию. Даже в отсутствие кольцевой структуры, линейные ДНК эукариотических клеток могут приобретать суперспирализованное состояние вследствие их взаимодействия с ядерными белками, что создает условия для формирования закрытых топологических доменов.
Можно с уверенностью утверждать, что явление отрицательной суперспирализации ДНК имеет важнейший биологический смысл, заключающийся в обеспечении компактной упаковки генетического материала, с одной стороны, и в облегчении функционирования процессов репликации и транскрипции ДНК, связанных с необходимостью незатрудненного разделения цепей в двухцепоченых молекулах ДНК, с другой стороны.