

|
|
|||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||
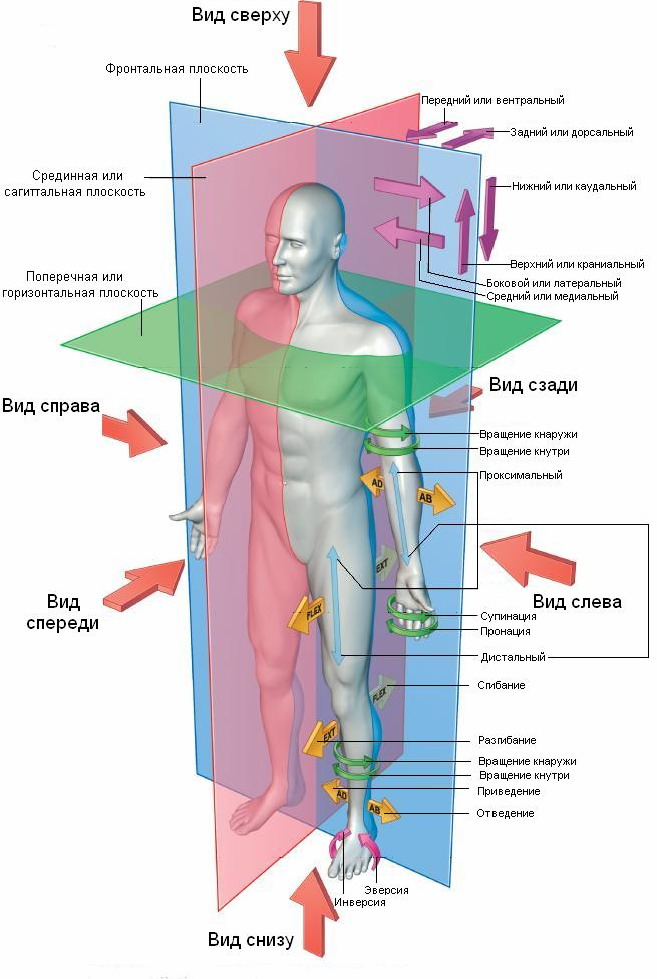
Рис. 1 Оси, плоскости и термины
-
ЧЕЛОВЕК. ОБЩИЕ ДАННЫЕ
В процессе изучения человека его структуры условно подразделяют на клетки, органы, системы и аппараты органов, которые и формируют организм. Однако следует предостеречь читателя от буквального понимания этого деления. Организм един, он может существовать лишь благодаря своей целостности.
КЛЕТКИ
Основной структурной единицей строения живого является клетка (cellula). Немецкий ученый Т. Шванн создал клеточную теорию, основные положения которой свидетельствуют о том, что все ткани состоят из клеток. Клетки растений и животных принципиально сходны между собой, все они возникают одинаково. Деятельность организма - сумма жизнедеятельности отдельных клеток.
Большое влияние на дальнейшее развитие клеточной теории и учение о клетке оказал Р. Вирхов. Он не только свел воедино все многочисленные разрозненные факты, но и убедительно показал, что клетки являются постоянной структурой и возникают только путем деления исходной клетки.
Клетка - элементарная единица всего живого, поэтому ей присущи свойства живых организмов: высокоупорядоченное строение; способность получать энергию извне и использовать ее для выполнения работы и поддержания функций; обмен веществ; способность активно реагировать на раздражение; рост; развитие; размножение; способность к самовоспроизведению и передаче генетической информации потомкам; регенерация, адаптация к окружающей среде.
Клеточная теория в современной интерпретации включает следующие главные положения: клетка является универсальной единицей живого; клетки всех организмов имеют принципиально сходное строение; клетки размножаются только путем деления исходной клетки; многоклеточные организмы являются сложными клеточными ансамблями, образующими целостные системы. Благодаря современным исследованиям выявлены два основных типа клеток: более сложно организованные высокодифференцированные эукариотические клетки (растения, животные и некоторые протисты - простейшие, водоросли, грибы и лишайники) и менее сложно организованные прокариотические клетки (синезеленые водоросли, актиномицеты, бактерии, спирохеты, микоплазмы, риккетсии, хламидии). В отличие от прокариотической эукариотическая клетка имеет ядро, ограниченное двойной ядерной мембраной, и большое количество мембранных органелл.
С точки зрения морфологии клетка представляет собой сложную систему биополимеров, отделенную от внешней среды клеточной мембраной (цитолеммой, плазмалеммой) и состоящую из ядра и цитоплазмы, в которой располагаются органеллы и включения (рис. 2).
Цитоплазма включает в себя гиалоплазму и органеллы общего назначения, которые имеются во всех клетках, и органеллы специального назначения, которые есть лишь в определенных клетках и выполняют специальные функции. В клетках встречаются также временные клеточные структуры – включения (рис.2). Клетки разнообразны по форме, строению, химическому составу и характеру обмена веществ. Все клетки гомологичны, т. е. имеют ряд общих структурных признаков, от которых зависит выполнение основных функций, присущих всем клеткам. Вместе с тем различные клетки имеют и специфические структуры. Это связано с выполнением ими специальных функций.
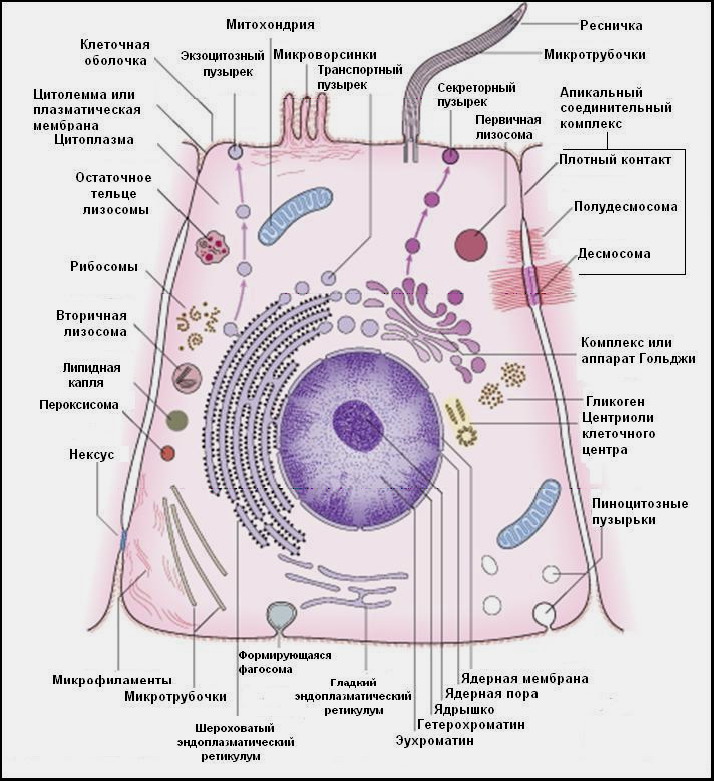
Рис.2 Ультрамикроскопическое строение клетки
Размеры клеток человека варьируют от нескольких микрометров (малый лимфоцит) до 200 мкм (яйцеклетка). В организме человека встречаются клетки различной формы: овоидные, шаровидные, веретеновидные, плоские, кубические, призматические, полигональные, пирамидальные, звездчатые, чешуйчатые, отростчатые и др.
Снаружи каждая клетка покрыта клеточной мембраной (цитолеммой, плазмалеммой) толщиной 9-10 нм, ограничивающей клетку от внеклеточной среды. Цитолемма выполняет следующие функции: защитную, разграничительную, рецепторную (восприятие сигналов внешней для клетки среды), транспортную, обеспечивает поверхностные свойства клетки.
Будучи очень тонкой, плазмалемма не видна в световом микроскопе. В электронном микроскопе на поперечном срезе последняя представляет собой трехслойную структуру, наружная поверхность которой покрыта тонкофибриллярным гликокаликсом толщиной от 7,5 до 200 нм. Гликокаликс состоит из боковых углеводных цепей гликолипидов и гликопротеидов, адсорбированных гликопротеидов и протеогликанов. Толщина наружного и внутреннего электронноплотных слоев (пластинок) мембраны составляет около 2,5 нм, среднего электронно-прозрачного слоя - около 3 нм. Однако часто на электронограммах мембрана выглядит однослойной.
Согласно жuдкостно-мозаuчной модели, разработанной в 1972 г. Г. Николсоном и С. Сингером, цитолемма, как и другие мембранные структуры, состоит из двух слоев амфипатическиx молекул липидов, лежащих перпендикулярно поверхности мембраны. При этом их гидрофильные «головки» направлены к наружной и внутренней сторонам мембраны, а гидрофобные «хвосты» обращены друг к другу (рис. 3).
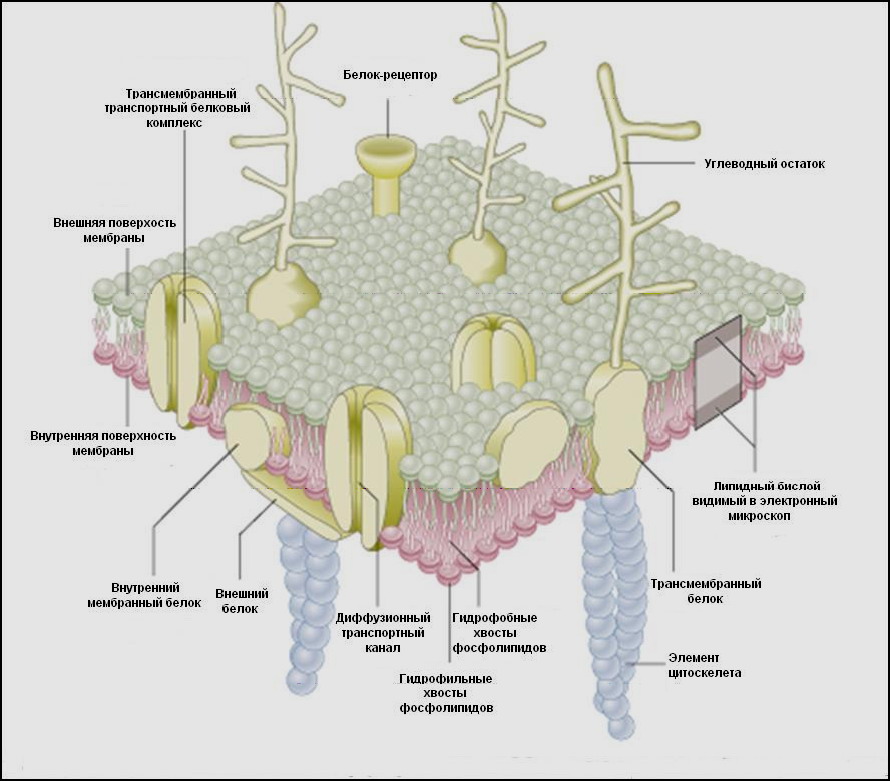
Рис. 3 Строение плазматической мембраны
В билипидный слой погружены молекулы белка. Некоторые из них (интегральные, или внутренние, трансмембранные) проходят через всю толщу мембраны, другие (периферические, или внешние) лежат во внутреннем или наружном монослое мембраны.
Некоторые интегральные белки связаны нековалентными связями с белками цитоплазмы. Подобно липидам, белковые молекулы также являются амфипатическими - их гидрофобные участки окружены аналогичными «хвостами» липидов, а гидрофильные обращены наружу или внутрь клетки.
Белки осуществляют большую часть мембранных функций: одни мембранные белки являются рецепторами, другие - ферментами, третьи - переносчиками.
Бислой асимметричен - в каждом монослое располагаются различные липиды. Гликолипиды обнаруживаются только в наружном монослое, причем их углеводные цепи направлены наружу. Молекулы холестерина в мембранах эукариот (у бактерий отсутствуют) лежат во внутренней, обращенной к цитоплазме половине мембраны. Цитохром с располагается в наружном монослое, а АТФ-синтетаза - на внутренней стороне мембраны.
Плазмалемма образует ряд специфических структур. Это межклеточные соединения, микроворсинки, реснички, клеточные инвагинации и отростки. Межклеточные соединения (контакты) подразделяются на простые и сложные. В простых соединениях плазмалеммы соседних клеток формируют выросты наподобие зубцов, так, что зубец одной клетки внедряется между двумя зубцами другой (зубчатое соединение), или переплетающихся между собой интердигитаций (пальцевидное соединение). Между плазмалеммами соседних клеток всегда сохраняется межклеточная щель шириной 15 - 20 нм.
Сложные контакты, в свою очередь, подразделяются на адгезионные, замыкающие и проводящие контакты. К адгезионным контактам относятся десмосома, полудесмосома и поясок сцепления (лентовидная десмосома). Десмосома состоит из двух электронно-плотных половин, которые принадлежат плазмалеммам соседних клеток, разделенных межклеточным пространством шириной около 25 нм, заполненным тонкофибриллярным веществом гликопротеидной природы. К обращенным к цитоплазме сторонам обеих пластинок десмосомы прикрепляются кератиновые тонофиламенты, напоминающие по форме головные шпильки. Помимо этого через межклеточное пространство проходят межклеточные волокна, соединяющие обе пластинки. Полудесмосома, образованная лишь одной пластинкой с входящими в нее тонофиламентами, прикрепляет клетку к базальной мембране. Поясок сцепления, или лентовидная десмосома, представляет собой «ленту», которая огибает всю поверхность клетки вблизи ее апикального отдела. Ширина межклеточного пространства, заполненного волокнистым веществом, не превышает 15 - 20 нм. Цитоплазматическая поверхность «ленты» уплотнена и укреплена сократительным пучком актиновых филаментов.
Плотные контакты проходят через апикальные поверхности клеток в виде поясков шириной 0,5 - 0,6 мкм. В плотных контактах между плазмалеммами соседних клеток практически нет межклеточного пространства и гликокаликса, а белковые молекулы обеих мембран контактируют между собой, поэтому через плотные контакты молекулы не проходят. На плазмалемме одной клетки имеется сеть гребешков, образованных цепочками эллиптических белковых частиц, расположенных во внутреннем слое мембраны. На плазмалемме соседней клетки этим гребешкам соответствуют углубления, бороздки.
К проводящим контактам относятся нексус, или щелевидный контакт, и синапс. Через эти соединения из одной клетки в другую проходят водорастворимые малые молекулы. Такими контактами соединены очень многие клетки человека (и животных). В нексусе между плазмалеммами соседних клеток имеется пространство шириной 2 - 4 нм. Обе плазмалеммы соединены между собой коннексонами - полыми гексагональными белковыми структурами размерами около 9 нм, каждая из которых образована шестью белковыми субъединицами. Щелевые контакты играют важную роль в осуществлении функций клеток, обладающих выраженной электрической активностью, например кардиомиоцитов.
Микроворсинки - это лишенные органелл пальцевидные выросты клетки, покрытые плазмалеммой, длиной около 1 - 2 мкм и диаметром до 0,1 мкм. В центре микроворсинки проходит пучок, состоящий из 30 - 40 параллельных актиновых филаментов диаметром около 3 нм каждый.
Микроворсинки покрыты слоем тонковолокнистого гликокаликса
толщиной 7 - 20 нм. Некоторые эпителиоциты, например кишечные, имеют очень большое количество микроворсинок, образуя так называемую исчерченную каемку. Наряду с микроворсинками на поверхности некоторых клеток есть крупные микроворсинки стереоцuлии (например, волосковые сенсорные клетки вестибулярного и улиткового лабиринтов, микроворсинчатые эпителиоциты протока придатка яичка и др.). Подобно микроворсинкам, их структурную основу составляют пучки параллельных актиновых филаментов, скрепленных между собой белковыми сшивками. Длина стереоцилий достигает 5 - 7 мкм.
Реснички и жгутики выполняют функцию движения. До 250 ресничек длиной 5 - 15 мкм и диаметром 0,15 - 0,25 мкм покрывают апикальную поверхность реснитчатого эпителиоцита верхних дыхательных путей, маточных труб, семявыносящих канальцев.
Клеточный центр образован двумя центриолями (дuплосома), окруженными ободком плотного вещества (центросферой), участком цитоплазмы, от которой радиально отходят микротрубочки. Клеточный центр расположен либо вблизи ядра, либо вблизи формирующейся поверхности комплекса Гольджи. Обе центриоли диплосомы расположены под углом друг к другу. Каждая центриоль представляет собой цилиндр, стенка которого состоит из девяти триплетов микротрубочек длиной около 0,5 мкм и диаметром около 0,25 мкм.
Центриоли являются полуавтономными самореплицирующимися структурами, которые удваиваются в клеточном цикле. Сначала обе центриоли расходятся и перпендикулярно базальному концу материнской, возникает мелкая процентриоль, образованная девятью микротрубочками. Затем к каждой из них из тубулина путем самосборки формируются еще две. Не исключено, что, подобно митохондриям, центриоли содержат собственную ДНК. Центриоли участвуют в образовании базальных телец ресничек и жгутиков и в образовании митотического веретена.
Мuкротрубочки, имеющиеся в цитоплазме всех эукариотических клеток, представляют собой полые цилиндры различной длины диаметром. 20 - 30 нм. Стенка микротрубочки толщиной 68 нм состоит из 13 параллельных протофиламентов, образованных описанными димерами тубулина. Микротрубочки образуют цитоскелет и участвуют в транспорте веществ внутри клетки.
В цитоплазме происходит постоянная самосборка микротрубочек из тубулина. Цитоплазматические микротрубочки растут из перицентриолярного вещества, окружающего каждую центриоль. Последние и являются центрами организации микротрубочек.
Цитоскелет клетки представляет собой трехмерную сеть, в которой различные белковые нити связаны поперечными сшивками. При этом различные органеллы и растворимые белки ассоциированы с цитоскелетом. В образовании цитоскелета участвуют микротрубочки (им принадлежит главная роль), а также актиновые, миозиновые и промежуточные волокна (филаменты). Однако актиновые субъединицы и филаменты входят в состав большинства клеток, они связываются с другими белками, в результате чего образуются трехмерные сети, пучки. В немышечных клетках движение также осуществляется благодаря взаимодействию актина с миозином. Белковые промежуточные филаменты толщиной 8 - 10 нм в различных клетках имеют не-одинаковый химический состав и строение: кератиновые тонофиламенты - в эпителиальных клетках, виметиновые - в некоторых клетках соединительной ткани, нейрофиламенты - в нейронах. Следует подчеркнуть, что при делении клетки в дочерних клетках образуется цитоскелет, аналогичный,такому в материнской. Иными словами, информация о строении цитоскелета передается по наследству.
Мембранные органеллы. Транспорт через мембраны. Для эукариотических клеток характерно наличие огромного количества внутриклеточных мембран, образующих множество компартментов (от англ. compartment - отделение, купе), отличающихся друг от друга строением и функцией: цитозоль, ядро, эндоплазматический ретикулум, комплекс Гольджи, митохондрии, лизосомы, пероксисомы. Благодаря наличию таких элементов в клетке пространственно разобщены одновременно протекающие различные биохимические реакции. При этом в клетке имеются определенные пути, по которым перемещаются синтезированные вещества, идущие на построение мембран, а также пути, по которым эти вещества поступают в клетку.
Все мембранные органеллы построены из элементарных мембран, строение которых аналогично строению плазмалеммы. Напомним об асимметрии мембран. Наружный, обращенный к цитозолю слой любой мембранной органеллы идентичен внутреннему слою плазмалеммы, а внутренний, обращенный в полость органеллы слой аналогичен наружному слою цитолеммы.
Одна из важнейших функций плазматической мембраны - транспорт. Напомним, что обращенные друг к другу «хвосты» липидных молекул образуют гидрофобный слой, препятствующий проникновению полярных молекул. Поступление веществ в клетку или выход их наружу осуществляется с помощью четырех основных механизмов: диффузии, осмоса, активного транспорта и экзо- или эндоцитоза. Первые два процесса носят пассивный характер (не требуют затрат энергии), два последних - активные процессы, связанные с потреблением энергии.
Пассивный транспорт незаряженных молекул осуществляется по градиенту концентрации, транспорт заряженных молекул зависит от градиента концентрации Н+ и трансмембранной разности потенциалов, которые объединяются в трансмембранный электрохимический градиент Н+, или электрохимический протонный градиент. Как правило, внутренняя цитоплазматическая поверхность мембраны несет отрицательный заряд, что облегчает проникновение в клетку положительно заряженных ионов.
Диффузия (от лат. diffusio - распространение, растекание) - это переход ионов или малых молекул, вызванный их броуновским движением через мембраны, из зоны, где эти вещества находятся в более высокой концентрации, в зону с более низкой концентрацией (по диффузионному градиенту).
Вода поступает в клетку путем осмоса (от греч. osmos - толчок, давление). В настоящее время математически доказывается наличие в цитолемме временных пор, возникающих по мере необходимости.
Существуют специфические транспортные белки, встроенные в мембрану, способные переносить через нее небольшие полярные молекулы определенного вида без затраты энергии (Н. Christensen, 1975). Это так называемая облегченная диффузия. Таким процессом является вход глюкозы в гепатоциты, эритроциты и мышечные клетки по концентрационному градиенту.
При активном транспорте молекулы движутся через мембрану независимо от ее концентрационного градиента. Энергия для такого процесса может поставляться одним из двух способов. Во-первых, транспорт данного соединения может быть сопряжен с одновременным движением второго вещества, которое движется по своему концентрационному градиенту. Вторая молекула может двигаться в том же направлении, что и первая (симnорт), или в противоположном направлении (антипорт). Во-вторых, энергия может поставляться сопряженным гидролизом АТФ на поверхности белка-переносчика. Такое «устройство» называется насосом. Оно формируется белками, прошивающими всю толщу мембраны. В тканях млекопитающих обнаружено несколько основных систем активного транспорта: натриевый и кальциевый насосы, системы транспорта глюкозы и других сахаров, системы транспорта аминокислот.
Поглощение клетками различных макромолекул и частиц происходит путем эндоцитоза, выделение - экзоцитоза. При эндоцитозе плазматическая мембрана образует впячивания или выросты, которые отшнуровываются и превращаются в пузырьки и вакуоли.
Известны две разновидности эндоцитоза: фагоцитоз - поглощение твердых частиц (от греч. phagos - пожирающий, cytos - клетка), nиноцитоз - поглощение растворенных веществ (от греч. pino - пью). В большинстве случаев при эндоцитозе поглощаемое вещество связывается с рецептором, встроенным в мембрану. Этот тип эндоцитоза осуществляется участками мембраны, выстланными изнутри особым белком клатрином - окаймленные ямки и пузырьки. Макрофаги и полиморфно-ядерные нейтрофиллы осуществляют фагоцитоз, опосредуемый рецепторами (Т. Stossel, 1974; J. Goldstein е. а., 1979).
Экзоцитоз - процесс, обратный эндоцитозу. Этим способом различные материалы выводятся из клеток: из пищеварительных вакуолей удаляются оставшиеся непереваренными плотные частицы, а из секреторных клеток выводится их жидкий секрет.
Эндоnлазматическая сеть, или эндоnлазматический ретикулум (ЭР), представляет собой единую непрерывную полость, ограниченную мембраной толщиной около 60 нм, которая образует множество инвагинаций и складок. Поэтому на электронограммах эндоплазматическая сеть выглядит в виде множества трубочек, плоских или округлых цистерн, мембранных пузырьков. Различают два типа ЭР: шероховатый, или гранулярный, и гладкий, или агра-нулярный. Обращенная к цитозолю сторона шероховатого ЭР покрыта рибосомами в отличие от гладкого ЭР, лишенного рибосом. Поэтому в световом микроскопе гранулярный ЭР выглядит как базофильное вещество, дающее положительную окраску на РНК. Функция гранулярного ЭР - синтез белков на рибосомах и транспорт белков, гладкого - синтез и метаболизм углеводов и липидов. В клетках, специализирующихся на синтезе белков, гранулярный эндоплазматический ретикулум выглядит в виде параллельных фенестрированных, сообщающихся между собой и с перинуклеарным пространством ламеллярных структур, между которыми лежит множество свободных рибосом.
Гладкий ЭР на электронограммах выглядит в виде множества мелких трубочек, диаметр просвета которых около 50 нм, между трубочками часто расположены гранулы гликогена. Гладкий ЭР осуществляет синтез стероидных гормонов (клетки Лейдига, корковые эндокриноциты надпочечника), синтез гликогена, холестерола и детоксикацию (гепатоциты), синтез хлоридов, из которых образуется соляная кислота (париетальные экзокриноциты желез желудка). Являясь депо ионов кальция, ЭР участвует в мышечном сокращении (миосимпласт), отграничивает будущие тромбоциты в мегакариоцитах. Одной из важнейших функций ЭР является синтез мембранных белков и липидов для всех клеточных органелл.
Комплекс, или аппарат, Гольджи - КГ (внутренний сетчатый аппарат) - представляет собой совокупность цистерн, пузырьков, пластинок, трубочек, мешочков, ограниченных мембраной, в которых накапливаются и упаковываются синтезированные продукты. Эти продукты с помощью элементов аппарата Гольджи выводятся из клетки, кроме того, в них происходят синтез полисахаридов, образование белково-углеводных комплексов и ковалентная модификация переносимых макромолекул. В световом микроскопе при импрегнации серебром аппарат Гольджи выглядит в виде сеточки или системы канальцев и вакуолей. В плазматических клетках это светлая зона над ядром, окруженная гранулярным ЭР («светлый дворик»). Комплекс Гольджи имеется во всех клетках человека, кроме эритроцитов и роговых чешуек эпидермиса. В большинстве клеток он расположен вокруг или вблизи ядра, в экзокринных клетках - над ядром, в апикальной части. При электронно-микрокопических исследованиях в аппарате Гольджи выявляются три мембранных элемента: уплощенные мешочки (цистерны), пузырьки и вакуоли. КГ является трехмерной структурой чашеобразной формы, состоящей из нескольких (от одной до нескольких сотен) диктиосом (от греч. dictyon - сеть). Каждая диктиосома диаметром около 1 мкм содержит 4-8 (в среднем 6) лежащих параллельно уплощенных цистерн, пронизанных порами, с расширенными концами, от которых отщепляются вакуоли, содержащие секреторные продукты. Цистерны ассоциированы с множеством мембранных везикул диаметром около 50 - б5 им (в том числе и окаймленные), которые располагаются главным образом на обращенной к ядру стороне и по периферии диктиосомы, а также с более крупными секреторными гранулами диаметром от 66 до 100 нм, локализованными на противоположной стороне диктиосомы. Элементы комплекса Гольджи связаны между собой каналами (W. Whally, 1975).
Аппарат Гольджи поляризован структурно и биохимически (А. Novicoff, 1977). В нем различают две поверхности: выпуклую формирующуюся (или цис-сторону), обращенную к эндоплазматическому ретикулуму и по строению напоминающую его, и вогнутую зрелую (или транс-сторону), обращенную к плазматической мембране и морфологически сходную с плазмалеммой. Мембраны аппарата Гольджи образуются и поддерживаются гранулярным эндоплазматическим ретикулумом, в котором синтезируются мембранные компоненты. Они переносятся транспортными пузырьками, отпочковывающимися от промежуточных зон ретикулума к формирующейся поверхности диктиосомы, и сливаются с ней. От транс-стороны постоянно отпочковываются секреторные пузырьки, а мембраны цистерн постоянно обновляются, они поставляют клеточную мембрану, гликокаликс и синтезированные вещества к плазмалемме. Таким образом обеспечивается обновление плазматической мембраны (G. Palade, 1975). Одной из важнейших функций комплекса Гольджи является сортировка белков.
Лuзосомы были открыты в 1955 г. Ch. dе Duve. Это мембранные органеллы диаметром 0,4 - 0,5 мкм, содержащие около 50 видов различных гидролитических ферментов: протеазы, фосфолипазы, нуклеазы, гликозидазы, фосфатазы, в том числе кислую фосфатазу. Последняя является маркером лизосом. Лизосомные ферменты синтезируются на рибосомах гранулярного эндоплазматического ретикулума, откуда переносятся транспортными пузырьками в аппарат Гольджи, где они модифицируются, и от зрелой поверхности отпочковываются первичные лизосомы. Все лизосомы клетки формируют лизосомное пространство, в котором с помощью протонного насоса постоянно поддерживается кислая среда - рН колеблется в пределах 3,5 - 5,0. Мембраны лизосом устойчивы к заключенным в них ферментам и предохраняют цитоплазму от их действия. Это связано с особой конформацией молекул лизосомной мембраны, при которой их химические связи экранированы. Повреждение или нарушение проницаемости мембраны лизосомы приводит к активации ферментов и тяжелым повреждениям, клетки, вплоть до ее гибели.
Различают четыре функциональные формы лизосомы. Первичные лизосомы, отпочковавшиеся от комплекса Гольджи, сливаясь с эндосомой, образуют вторичную лuзосому (фаголизосому), в которой происходит переваривание биополимеров до мономеров. Последние транспортируются через лизосомную мембрану в цитозоль. Непереваренные вещества остаются в лизосоме, в результате чего образуется остаточное тельце, содержащее вещество высокой электронной плотности. Вторичные лизосомы идентифицируются благодаря наличию в них осмиофильного вещества различной электронной плотности.
Вторичные лизосомы могут сливаться между собой, а также с другими первичными лизосомами. При слиянии пиноцитозных пузырьков с первичными лизосомами иногда образуются своеобразные вторичные лизосомы - мультuвезuкулярные тельца.
В процессе жизнедеятельности клетки постоянно происходят процессы физиологической регенерации на разных иерархических уровнях, начиная от молекул и кончая органеллами. Вблизи поврежденных участков цитоплазмы различных органелл и их фрагментов, обычно по соседству с комплексом Гольджи, образуется полулунная двойная мембрана (из эндоплазматического ретикулума), которая растет, окружая со всех сторон поврежденные зоны. Затем эта структура сливается с лизосомами (аутолuзосома). В других случаях структуры (например, гранулы секрета) впячиваются в лизосомную мембрану, окружаются ею и подвергаются перевариванию. Образуется аутофагuческая вакуоль. В результате множественной микроаутофагии образуются мульти-везикулярные тельца (например, в нейронах мозга, кардиомиоцитах и др.).
Аутофагuя - переваривание участков собственного вещества («самопоедание») - происходит и при недостатке пищевых веществ. Наряду с аутофагией в некоторых клетках происходит и крuнофагuя (от греч. krinos - просеивать, отделять) - слияние первичных лизосом с секреторными гранулами. В лизосомах необновляющихся клеток в результате многократного аутофагирования накапливается липофусцин - пигмент старения.
Поступление в клетку твердых частиц (фагоцитоз) или капель жидкости (пиноцитоз) извне называется гетерофагuей (от греч. heteros - иной). Этот процесс наиболее широко распространен у простейших, но часто встречается у человека (и других млекопитающих). Гетерофагия играет важную роль в защите организма (сегментоядерные нейтрофильные гранулоциты, макрофагоциты), перестройке костной ткани (остеокласты), образовании тироксина фолликулами щитовидной железы, реабсорбции белка и других макромолекул в проксимальном отделе нефрона и других процессах.
Пероксuсомы представляют собой пузырьки диаметром от 0,2 до 0,5 мкм, которые окружены мембранами и содержат окислительные ферменты (около 40 % всех белков составляет каталаза), производящие и разрушающие пероксид водорода. Они используют молекулярный кислород.
Мuтохондрuu, являющиеся «энергетическими станциями клетки», участвуют в процессах клеточного дыхания и преобразования энергии в форму, доступную для использования клеткой. В световом микроскопе митохондрии выглядят в виде округлых, удлиненных или палочковидных структур длиной 0,3 - 5 мкм и шириной 0,2 - 1 мкм. Количество, размеры и расположение митохондрий зависят от функции клетки, ее потребности в энергии. Так, в гепатоцитах их количество достигает 2500.
С помощью электронной микроскопии установлено, что митохондрии являются органеллами с двойными мембранами (толщиной около 7 нм каждая). Между наружной и внутренней митохондриальными мембранами расположено межмембранное пространство толщиной 10 - 20 нм.
Внутренняя мембрана образует многочисленные складки, или кристы. Митохондрии (у растений также хлоропласты) содержат собственную ДНК (кольцевая митохондриальная хромосома, D. Clayton, 1982). Около 2 % ДНК клетки содержится в митохондриях. Митохондрии обладают РНК и рибосомами, которые также находятся в матриксе. Таким образом, митохондрии снабжены собственным аппаратом самовоспроизведения, синтеза белка (А. Tzagolof, 1979). Следует подчеркнуть, что митохондриальные ДНК, РНК и рибосомы отличаются от таковых собственной клетки, но сходны с этими структурами у прокариот. У млекопитающих (и человека) митохондриальный геном наследуется от матери (R. Gilles, 1980). Митохондрии (и хлоропласты) размножаются путем бинарного деления независимо от деления других митохондрий и самой клетки. Таким образом, митохондрии (и хлоропласты) являются полуавтономными органеллами.
Каждая структура митохондрий обладает собственным, только ей присущим набором ферментов, выполняет строго определенную функцию.
Клеточное ядро. Ядро - основная клеточная структура; имеется во всех клетках человека, кроме эритроцитов и тромбоцитов. У большинства клеток его форма шаровидная или овоидная, однако, встречаются и другие формы ядра (кольцевидное, палочковидное, веретеновцдное, четковидное, бобовидное, сегментированное, полиморфное и др.). Размеры ядер колеблются в широких пределах - от 3 до 25 мкм. Наиболее крупное ядро имеет яйцеклетка. Большинство клеток человека одноядерные, однако, имеются двухъядерные (например, некоторые нейроны, гепатоциты, кардиомиоциты), а некоторые структуры многоядерные (мышечные волокна - миосимпласты). В ядре различают следующие структуры: ядерную оболочку, хроматин, ядрышко и нуклеоплазму. Ядро окружено ядерной оболочкой, состоящей из внутренней и наружной ядерных мембран толщиной 8 нм каждая, разделенных перu-нуклеарным пространством (или цистерной ядерной оболочки) шириной 20-50 нм.
Обе являются элементарными клеточными мембранами. К наружной мембране, переходящей в гранулярный эндоплазматический ретикулум, прикреплены рибосомы. Перинуклеарное пространство составляет единую полость с эндоплазматическим ретикулумом. Ядерная оболочка пронизана множеством расположенных упорядоченно ядерных пор округлой формы диаметром 50-70 нм, которые в общей сложности занимают до 25 % поверхности ядра. По краям пор наружная или внутренняя ядерные мембраны соединяются одна с другой. Комплекс поры представляет собой сложную структуру, которая состоит из трех рядов связанных между собой белковых гранул (в каждом ряду по восемь гранул), располагающихся на равном расстоянии друг от друга по обе стороны ядерной оболочки, и одной гранулы в центре. Через ядерные поры осуществляется избирательный транспорт крупных частиц, а также обмен веществ между ядром и цитозолом.
В живых клетках кариоплазма (нуклеоплазма) гомогенна (кроме ядрышка). После фиксации и обработки тканей для световой или электронной микроскопии в кариоплазме становятся видными два типа хроматина (от греч. chroma - краска): хорошо окрашивающийся электронно-плотный гетерохроматин и светлый эухроматин. Гетерохроматин расположен в основном вблизи ядерной мембраны и вокруг ядрышка, эухроматин находится между скоплениями гетерохроматина. Хроматин - это хромосомный материал, экстрагируемый из ядер эукариотических клеток. В его состав входят ДНК, небольшие основные белки гистоны, более крупные кислые белки и небольшое количество РНК. В делящемся ядре хроматин спирализуется, конденсируется, в результате чего становятся видимыми хромосомы. Молекула ДНК состоит из двух правозакрученных полинуклеотидных цепей (двойная спираль), имеющих антипараллельное направление и связанных между собой водородными связями. Молекула РНК образована одной полинуклеотидной цепью.
Молекулы РНК и ДНК отличаются по составу азотистых оснований и сахара. ДНК содержит аденин (А), гуанин (G), цитозин (С) и тимин (Т), сахар-дезоксирибозу, РНК-аденин, гуанин, цитозин и урацил (U), сахар-рибозу. Каждая хромосома состоит из одной молекулы ДНК. Ген - участок ДНК, характеризующийся определенной последовательностью нуклеотидов, ответственный за синтез одного определенного специфического белка. Ген является элементарной единицей наследственности.
Молекула ДНК упакована компактно.
Длина одной хромосомы человека в растянутом виде составляет около 5 см (В. Албертс и др., 1986).
ДНК ассоциирована с гистонами - белками основной природы, в результате чего образуются нуклеосомы, являющиеся структурными единицами хроматина.
Нуклеосомы, напоминающие бусины, состоят из восьми молекул гистонов, вокруг которых закручен участок ДНК, включающий 146 пар нуклеотидов. В результате последующей укладки метафазная хроматида укорочена в 8000 раз по сравнению с молекулой ДНК.
В 1956 г. F. Tjio, А. Levan открыли, что соматические (диплоидные) клетки человека содержат 46 хромосом, половые (гаплоидные) - 23. С помощью электронной микроскопии была выявлена структура хромосом. Метафазная хромосома состоит из двух соединенных центромерой сестринских хроматид, каждая из которых содержит одну молекулу ДНК, уложенную в виде суперспирали. Поверхность хромосомы покрыта различными молекулами, главным образом РНК.
Ядрышко (одно или несколько) выявляется во всех неделящихся ядрах в виде плотного интенсивно окрашивающегося округлого однородного базофильного тельца, размеры тем больше, чем интенсивнее протекает белковый синтез. Размеры ядрышка зависят в основном от количества гранулярного компонента, которое контролируется на уровне транскрипции рибосомных генов (S. Ghosh, 1976). Ядрышко состоит из 'двух отличающихся друг от друга компонентов: электронно-плотной нуклеолонемы (от греч. nema - нить), в которой различают нитчатую (фибриллярную) часть, состоящую из рибонуклеопротеинов (РНП) и множества переплетающихся нитей РНК, и гранулярную часть, образованную зернами - частицами РНП, предшественниками рибосомных субъединиц. В ядрышке образуются рибосомы.
Нуклеоплазма - неокрашивающаяся электронно-прозрачная часть ядра - коллоидный раствор белков, окружающий хроматин и ядрышко.
Цитозоль. Рибосомы и синтез белка. Цитозоль представляет собой часть цитоплазмы, окружающей органеллы, занимает около 53-55 % общего объема клетки (В. Албертс и др., 1986). В нем содержится огромное количество ферментов, катализирующих различные реакции промежуточного обмена, а также белки цитоскелета. На электронограммах цитозоль выглядит в виде аморфного вещества малой электронной плотности. На расположенных в нем рибосомах синтезируются многие белки.
Рибосомы представляют собой округлые осмиофильные структуры размерами 20 Х 30 нм, состоящие из двух субъединиц: большой, в состав которой входят три различные молекулы РНК, связанные с 40 молекулами белка, и малой, содержащей одну молекулу рРНК и около 33 молекул белка (В. Албертс и др., 1986).
Рибосомы, осуществляющие синтез белка, присутствуют во всех клетках человека, за исключением зрелых эритроцитов. Между обеими субъединицами рибосомы имеется щель, в которой проходит молекула информационной РНК, на рибосоме - бороздка, где располагается полипептидная цепь. Соотношение рРНК: белок составляет примерно 1:1. Именно рибосомы и полирибосомы обусловливают базофилию некоторых участков цитоплазмы. Рибосомы расположены поодиночке (монорибосомы) или группами в виде розеток, спиралей, завитков (полирибосомы, или полисомы). Последние образуются в результате связывания нескольких рибосом с одной молекулой мРНК.
Рибосомы и полисомы могут свободно располагаться в цитоплазме или, как это указывалось ранее, прикрепляться к мембранам гранулярного эндоплазматического ретикулума. Свободные рибосомы синтезируют белок, необходимый для жизнедеятельности самой клетки, прикрепленные - белок, подлежащий выведению из клетки.
Синтез белка (трансляция) связан с процессом транскрипции - переписывания информации, хранящейся в ДНК. Информация о структуре белка, заключенная в ДНК, «переписывается» на информационную, или мессенджер-РНК (мРНК) (от англ. messenger - посредник). Небольшие транспортные РНК (тРНК) выполняют двойную функцию: они присоединяют молекулу аминокислоты, транспортируют ее в рибосому и узнают триплет (три нуклеотида), соответствующий этой аминокислоте в молекуле мРНК.
Клеточный цикл (деление клетки). Клеточный цикл представляет собой совокупность процессов, происходящих в клетке при подготовке ее к делению и во время деления, в результате которого материнская клетка делится на две дочерние.
В клеточном цикле выделяют две фазы: автосинтетическую (подготовка клетки к делению) и митоз.
Соматические клетки содержат по две копии каждой хромосомы, их называют гомологичными. Они одинаковы по длине; форме, строению, расположению полос и несут одни и те же гены, которые локализованы одинаково. Нормальный кариотип человека (от греч. karion - ядро ореха, typos - образец) включает 22 пары аутосом и одну пару половых хромосом (ХХ или ХУ) (рис. 4).
Клеточный цикл подразделяется на митоз и интерфазу, которая представляет собой промежуток времени между окончанием одного митоза и началом следующего.
Митоз (от греч. mitos - нить) подразделяется на профазу, метафазу, анафазу и телофазу (рис. 5).
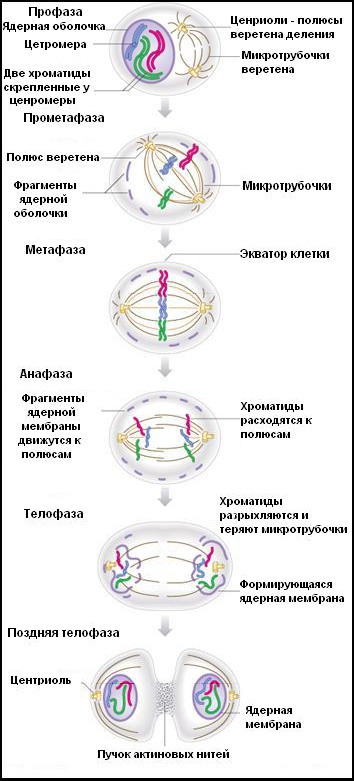
Рис. 5 Стадии митоза
Митоз обеспечивает генетическую стабильность, увеличение числа клеток в организме и, следовательно, рост организма, а также процесс регенерации.
Мейоз - своеобразный тип деления клеток, при котором происходит уменьшение числа хромосом. Это деление называют также редукционным делением клеток. Такое деление обязательно проходят половые клетки, у которых в процессе созревания в результате мейоза образуется не только уменьшенное вдвое (гаплоидное) число хромосом, но и новая комбинация генов в хромосомах.
Поэтому новый организм несет не просто сумму черт своих родителей, а является новым образованием, с присущими только ему признаками.
При мейозе происходят два последовательных (мейотических) клеточных деления. Вначале в исходных (первичных) половых клетках (ооцитах или сперматоцитах первого порядка - оогониях или сперматогониях), имеющих удвоенное (диплоидное) число хромосом (46) , происходит внутрихромосомная рекомбинация, когда гомологичные родительские хромосомы, имеющие либо материнское, либо отцовское происхождение, обмениваются своими идентичными участками. В результате этого процесса, который называется кроссинговером, и клеточного деления каждая дочерняя клетка имеет гаплоидное число хромосом (23) и наследственные признаки обоих родителей. У особей женского пола в результате двух мейотических делений первичной половой клетки образуются четыре дочерние клетки. Из них только одна становится зрелой половой клеткой с гаплоидным числом хромосом (23), а остальные три образуют так называемые полярные (направительные) тельца, в дальнейшем дегенерирующие. У особей мужского пола из первичной половой клетки (сперматогония) также образуются четыре дочерние клетки, имеющие по одному (гаплоидному) числу хромосом (23). Все эти четыре клетки развиваются в зрелые сперматозоиды.
-
УЧЕНИЕ О КОСТЯХ (ОСТЕОЛОГИЯ)
Одной из важнейших функций организма человека является передвижение в пространстве. Ее выполняет опорно-двигательный аппарат, состоящий из двух частей: пассивной и активной. К первой относятся кости, соединяющиеся при помощи различного вида соединений, ко второй - мышцы.
Скелет (от греч. skeletos - высохший, высушенный) представляет собой комплекс костей, выполняющих множество функций: опорную, защитную, локомоторную, формообразующую, преодоление силы тяжести. Форма тела человека обусловлена скелетом, имеющим билатеральную симметрию и сегментарное строение (рис. 6). Общая масса скелета составляет от 1/7 до 1/5 массы тела человека. В состав скелета человека входит более 200 костей. 33-34 кости скелета - непарные. Это позвонки, крестец, копчик, некоторые кости черепа и грудина, остальные кости парные. Скелет условно подразделяют на две части: осевой и добавочный. К осевому скелету относятся: позвоночный столб (26 костей), череп (29 костей), грудная клетка (25 костей). К добавочному - кости верхних (64) и нижних (62) конечностей.
Кости скелета являются рычагами, приводимыми в движение мышцами. В результате этого части тела изменяют положение по отношению друг к другу и передвигают тело в пространстве. К костям прикрепляются связки, мышцы, сухожилия, фасции, являющиеся элементами мягкого остова или мягкого скелета, который также принимает участие в удерживании органов возле костей, образующих твердый (жесткий) скелет.
Скелет образует вместилище для органов, защищая их от внешних воздействий: в полости черепа расположен головной мозг, в позвоночном канале - спинной мозг, в грудной клетке - сердце и крупные сосуды, легкие, пищевод и др., в полости таза - мочеполовые органы.
Кости участвуют в минеральном обмене, они являются депо кальция, фосфора и т. д. Живая кость содержит витамины А, D, С и др. Жизнедеятельность кости зависит от функций гипофиза, щитовидной и паращитовидных желез, надпочечников и гонад.
Скелет образован разновидностями соединительной ткани - костной и хрящевой, которые состоят из клеток и плотного межклеточного вещества. Кость и хрящ тесно связаны между собой общностью строения, происхождения и функции. Большинство костей развивается из хряща, их рост обеспечивается за счет его пролиферации (кости конечностей, основания черепа, позвонки).
Небольшое количество костей развивается без участия хряща (кости крыши черепа, нижняя челюсть, ключица). Некоторые хрящи не связаны с костью и в течение всей жизни человека не заменяются ею (хрящи ушных раковин, воздухоносных путей). Некоторые хрящи связаны с костью функционально (суставные хрящи, мениски, суставные губы). У зародыша человека и других позвоночных животных хрящевой скелет составляет около 50 % массы всего тела. Однако постепенно хрящ заменяется костью, у взрослого человека масса хряща достигает около 2 % массы тела. Это суставные хрящи, межпозвоночные диски, хрящи носа и уха, гортани, трахеи, бронхов и ребер. Хрящи выполняют следующие функции:
Покрывают сочленованные поверхности, обладающие благодаря этому высокой устойчивостью к износу.
Суставные хрящи и межпозвоночные диски, являющиеся объектами приложения, сил сжатия и растяжения, осуществляют их передачу и амортизацию.
Хрящи воздухоносных путей и наружного уха формируют стенки полостей. К другим хрящам прикрепляются мышцы, связки, сухожилия.
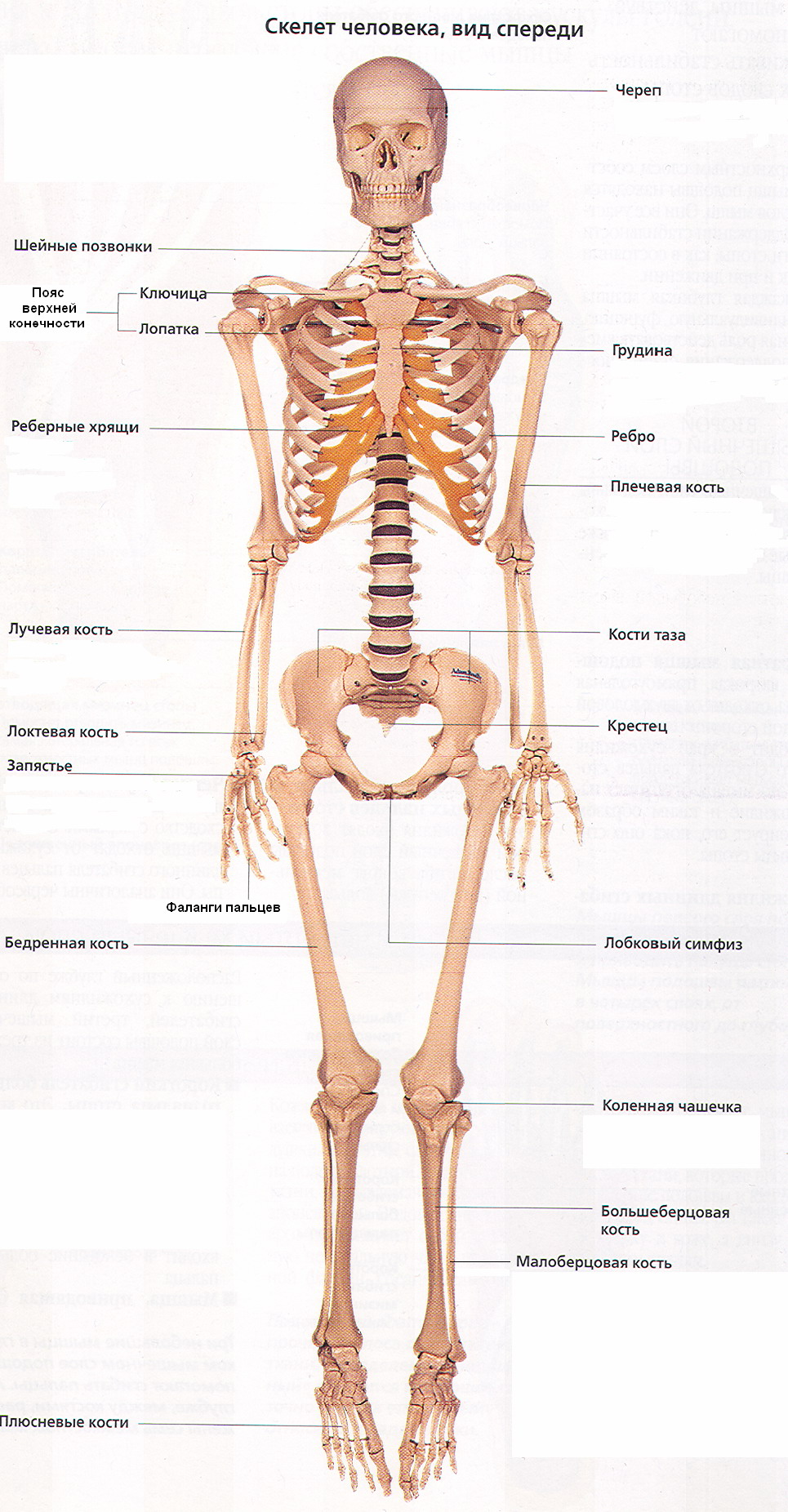
Рис 6 а
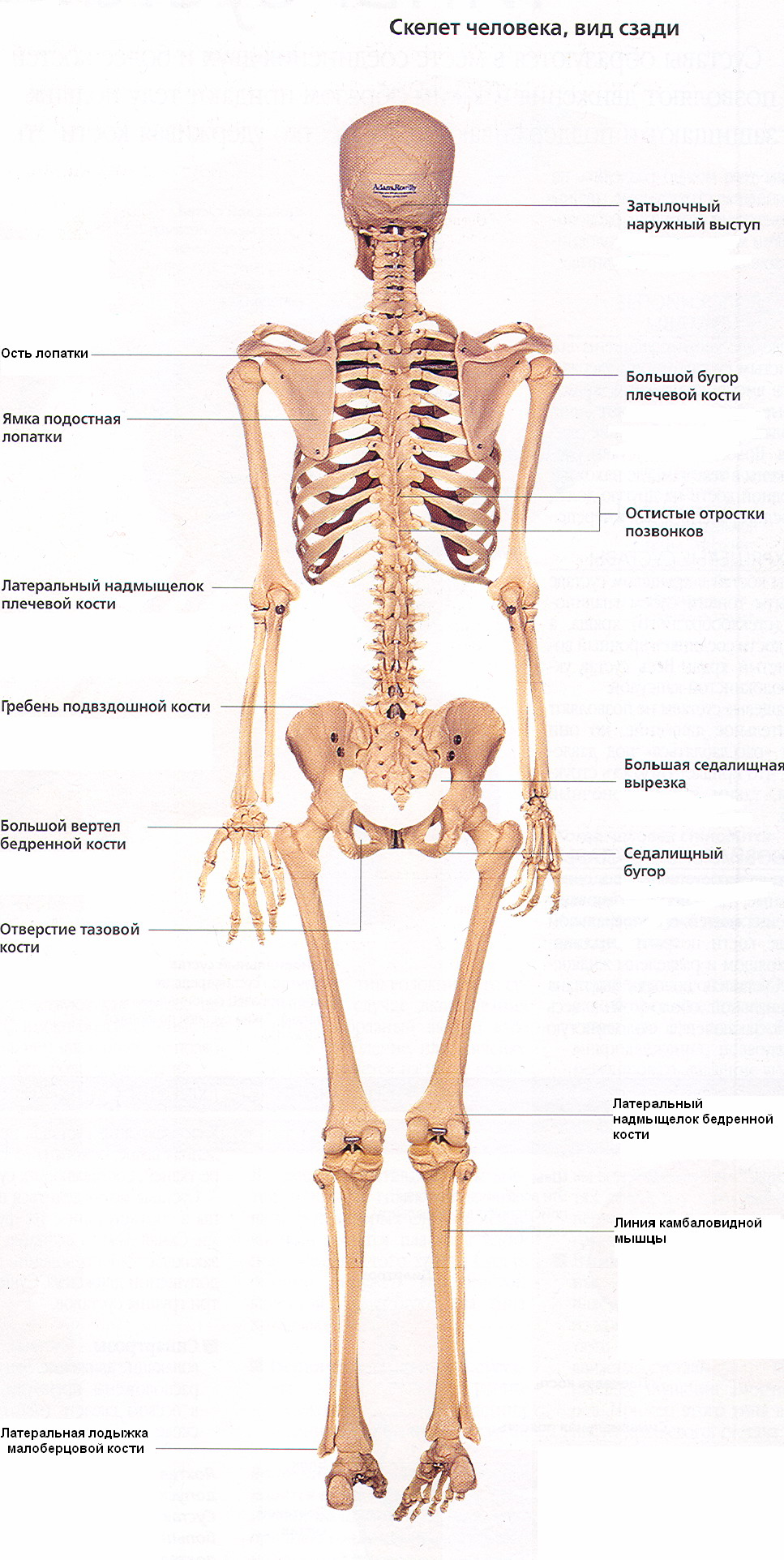
Рис 6 б
Хрящевая ткань содержит около 70-80 % воды, 10-15% органических веществ, 4 - 7 % солей. Около 50-70 % сухого вещества хряща приходится на долю коллагена. В зависимости от состава хрящи бывают гиалиновые, эластические и коллагеноволокнистые. Подобно другим разновидностям соединительной ткани, хрящевая ткань состоит из немногочисленных клеток и вырабатываемого ими плотного межклеточного вещества (матрикса). В зрелом гиалиновом хряще матрикс образован тремя компонентами: петлистым каркасом, образованным коллагеном II типа, ячейки которого заполнены молекулами протеогликанов и интерстициальной жидкостью, циркулирующей в этих пространствах в соотношениях 2,5:6,5.
Основные специализированные клетки хрящевой ткани, которые вырабатывают все компоненты хрящевого матрикса одновременно, - хондроциты. Крупные хондроциты имеют округлую, овальную или полигональную форму, базофильную цитоплазму, богатую элементами гладкого и шероховатого эндоплазматического ретикулума. В них хорошо развит комплекс Гольджи, содержащий много гранул, окрашивающихся рутениевым красным, а также имеется множество включений гликогена и липидов. Макромолекулы протеогликанов синтезируются клеткой, а их самосборка в агрегаты и образование комплексов с коллагеном происходят в матриксе. Ядро хондроцита, расположенное в центре или эксцентрично, богато хроматином. Цитолемма образует множество микроворсинок, которые контактируют с матриксом. Однако коллагеновые и эластические волокна непосредственно не соприкасаются с цитолеммой и находятся от нее на расстоянии 1-2 мкм.
Хондроциты, окруженные межклеточным веществом, располагаются в полостяхлакунах (рис. 7) . Если в лакуне содержится одна клетка, лакуна называется первичной. Чаще всего клетки располагаются в виде изогенных групп, которые представляют собой клеточные клоны. В таких случаях изогенная группа, или клеточное гнездо, лежит в первичной лакуне, а каждая из клеток этой группы расположена в своей собственной вторичной лакуне.
.

Структурной и функциональной единицей хряща является хондрон, образованный клеткой или изогенной группой клеток, околоклеточным - матриксом и капсулой лакуны.
Хрящи не имеют кровеносных сосудов, их питание осуществляется за счет диффузии из окружающих тканей. Хрящ как орган построен из хрящевой ткани, покрытой надхрящницей, состоящей из двух слоев: наружного, образованного волокнистой соединительной тканью, богато васкуляризированной и иннервированной, и внутреннего, хондрогенного, в котором лежат хондробласты и прехондробласты. Непосредственно под надхрящницей находятся хондроциты веретенообразной формы, располагающиеся несколькими слоями параллельно надхрящнице. Эти клетки весьма напоминают по своему строению фибробласты. Глубже располагаются постепенно укрупняющиеся и округляющиеся хондроциты. Еще глубже в лакунах лежат изогенные группы. В некоторых хрящевых клетках можно увидеть фигуры митоза.
Гиалиновый хрящ гладкий, блестящий, голубовато-белого цвета. Строение его матрикса описано выше. Следует указать, что коллагеновые волокна проходят через весь хрящ от одного листка надхрящницы к противоположному. Из гиалинового хряща образован в основном скелет эмбриона, у взрослого человека - реберные хрящи, большинство хрящей гортани, хрящи носа, трахеи, бронхов и суставные (с возрастом гиалиновый хрящ кальцифицируется).
Эластический хрящ менее прозрачен, желтоватого цвета. Строение его напоминает строение гиалинового хряща, однако в эластическом хряще количество изогенных групп меньше. Особенностью эластического хряща является наличие в его матриксе большого количества эластических волокон, образующих плотную сеть. Стенки лакун также сформированы эластическими волокнами. Ушная раковина, хрящевой отдел наружного слухового прохода, надгортанник, голосовые отростки черпаловидных хрящей гортани и слуховая труба состоят из эластической хрящевой ткани.
Волокнистый хрящ образует межпозвоночные диски, мениски, коленного и височно-нижнечелюстного суставов, лобковый симфиз. Он также имеется в зонах прикрепления связок и сухожилий к костям и хрящам. В отличие от описанных выше типов хряща в матриксе волокнистого большая часть коллагена (около 90 %)это коллаген I типа, содержание коллагена II типа очень небольшое; коллагеновые волокна расположены упорядоченными параллельными рядами, которые хорошо видны. Содержание хондроцитов в волокнистом хряще значительно ниже, чем в гиалиновоми эластическом хрящах, расположены они также в лакунах, реже встречаются изогенные группы. Клетки суставных дисков и менисков не синтезируют хондроитинсульфатов, не лежат в лакунах и лишены капсул.
Протеогликаны удерживают воду в матриксе хряща, поэтому прочность хряща на сжатие коррелирует с содержанием протеогликанов в его матриксе. Чем больше протеогликанов и меньше воды, тем выше жесткость хряща; иными словами, чем больше протеогликанов в матриксе хряща, тем меньшее количество жидкости удаляется из хряща при компрессии. Согласно современной биомеханической модели, матрикс хряща состоит из жидкой (воды) и твердой (протеогликаны и коллаген) фаз. Под влиянием компрессии происходит обратимая деформация хряща.
Вначале в связи с вытеснением воды из матрикса имеет место крутой подъем кривой, после чего дальнейшая деформация связана с изменениями твердой фазы. В дальнейшем наступает динамическое равновесие.
Кости образованы костной тканью, механические свойства которой обусловливают функции костей. Так, сопротивление свежей кости и чистой меди на растяжение одинаковы и в 9 раз больше, чем сопротивление свинца. Кость выдерживает сжатие10 кг/мм2 (аналогично чугуну), в то время как кирпич – лишь 0,5 кг/мм2. Предел прочности ребер на излом составляет 110 кг/см2.
Это связано с особенностями химического состава, структуры и архитектоники костей. Содержание воды в кости достигает 50 %. В сухом остатке костной ткани содержится около 33 % органических и 67 % неорганических веществ.
Подобно хрящу, кость состоит из клеток и межклеточного матрикса. Различают костные клетки двух типов: остеобласты и остеоциты (рис. 8).
8).
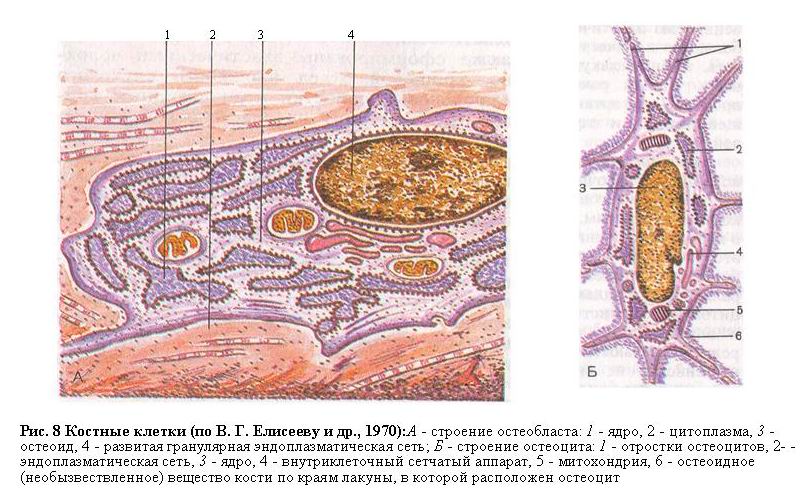
Остеобласты - это многоугольные, кубические, отростчатые молодые клетки с резко базофильной цитоплазмой, хорошо развитыми зернистой цитоплазматической сетью и комплексом Гольджи, с большим количеством рибосом. Многочисленные отростки остеобластов, богатые актиноподобными микрофиламентами, контактируют между собой и с отростками остеоцитов. Остеобласты залегают главным образом в поверхностных слоях кости. Округлое или овальное ядро богато хроматином и содержит одно крупное ядрышко, обычно расположенное на периферии. Расширенные полости эндоплазматического ретикулума содержат осмиофильное вещество. Остеобласт окружен тонкими коллагеновыми микрофибриллами, образующими микроокружение клетки, лишенное солей кальция. Матрикс, пропитанный кристаллами гидроксиапатита, расположен на расстоянии 12 мкм от плазмалеммы остеобласта.
Остеобласты синтезируют компоненты межклеточного вещества (проколлаген, сульфатированные гликозаминогликаны, протеогликаны) и выделяют их из клетки через всю поверхность в разных направлениях, что и приводит к образованию лакун, в которых они залегают, превращаясь в остеоциты. Наиболее активно синтезируется коллаген I типа, который во внеклеточном пространстве собирается в рыхло лежащие коллагеновые волокна.
В промежутке между ними располагается аморфное вещество, состоящее из протеогликанов, сульфатированных гликозаминогликанов, органических кислот. Этот комплекс представляет собой остеоидную ткань, или предкость, которая кальцифицируется. Органический матрикс кости импрегнируется кристаллами гидроксиапатита Ca10(PО4)6(OH)2 и аморфным фосфатом кальция Са3(РО4)2, которые поступают в костную ткань из крови через тканевую жидкость. Кристаллы гидроксиапатита в виде слоя окружают коллагеновые фибриллы, а также присутствуют внутри фибрилл. Мостики, образованные кристаллами, соединяют фибриллы между собой. Кристаллы импрегнируют и аморфное вещество.
Остеоциты - зрелые многоотростчатые веретенообразные клетки с крупным округлым ядром, в котором четко видно ядрышко. Количество органелл невелико - это митохондрии, элементы шероховатого эндоплазматического ретикулума и комплекса Гольджи. Остеоциты располагаются между костными - пластинками в лакунах, однако тела клеток не соприкасаются непосредственно с кальцифицированным матриксом, будучи окаймленными тонким слоем (1-2 мкм) остеоидной (неминерализованной) ткани, образованной коллагеновыми микрофибриллами и аморфным веществом. Очень длинные (до 50 мкм) отростки остеоцитов, богатые актиноподобными микрофиламентами, проходят в канальцах, причем они отделены от кальцифицированного матрикса пространством шириной около 0,1 мкм, в котором циркулирует тканевая жидкость, осуществляющая трофику клеток. Расстояние между каждым остеоцитом и ближайшим капилляром не превышает 0,1-0,2 мм.
В костной ткани имеется еще одна категория клеток - остеокласты, которые не являются костными, а имеют моноцитарное происхождение и относятся к системе макрофагов. Остеокласты - это крупные многоядерные (5-100 ядер) клетки, размерами до 190 мкм, которые разрушают кость и хрящ. Ядра богаты хроматином и имеют хорошо видимые ядрышки. В цитоплазме содержатся множество митохондрий, элементы шероховатого эндоплазматического ретикулума и комплекса Гольджи, свободные рибосомы, различные функциональные формы лизосом.
Остеокласты имеют огромное количество цитоплазматических отростков, которых особенно много на поверхности, прилежащей к разрушаемой кости, - гофрированная каемка, увеличивающая площадь соприкосновения клетки с костью. Отростки, в свою очередь, покрыты микроворсинками, между которыми находятся кристаллы гидроксиапатита. Кристаллы обнаруживаются также в фаголизосомах клетки, где они подвергаются фагоцитозу. Благодаря разрушению кости образуются лакуны Хаушипа, в которых залегают остеокласты. На поверхности клетки, там, где она соприкасается с костью, видна щеточная каемка, которая образована множеством обнаженных коллагеновых фибрилл после того, как неорганический компонент матрикса разрушен. Фибриллы проникают между ворсинками гофрированной каемки. Последняя окружена прилежащей к кости светлой зоной, очень бедной органеллами, но содержащей актиноподобные филаменты. Ближе к центру клетки располагается зона пузырьков и вакуолей, часть из них представляет собой углубления между ворсинками гофрированной каемки. Вокруг ядра находится базальная область цитоплазмы, где расположено большинство органелл.
Различают два типа костной ткани: ретuкулофuброзную (грубоволокнuстую) и пластинчатую. Первая имеется у зародыша человека, у взрослго она располагается в зонах прикрепления сухожилий к костя, в швах черепа после их зарастания. Ретикулофиброзная костная ткань состоит из толстых неупорядоченных пучков коллагеновых волокон, между которыми находится аморфное вещество. В лакунах залегают остеоциты, их строение и расположение описаны выше. Грубоволокнистая кость снаружи покрыта надкостницей.
Пластuнчатая костная ткань наиболее распространена в организме. Она образована костными пластинками толщиной от 4 до 15 мкм, которые состоят из остеоцитов и тонковолокнистого костного основного вещества.
Волокна, образующие пластинки, лежат параллельно друг другу и ориентированы в определенном направлении. При этом волокна соседних пластинок разнонаправлены и перекрещиваются почти под прямым углом, что обеспечивает большую прочность кости.
Кость снаружи, кроме сочленованных поверхностей, покрыта надкостницей, представляющей собой прочную соединительнотканную пластинку, богатую кровеносными и лимфатическими сосудами, нервами. Надкостница прочно сращена с костью при помощи соединительнотканных прободающих (шарпеевских) волокон, проникающих в глубь кости. Наружный слой надкостницы волокнистый, фиброзный, образован коллагеновыми волокнами, среди которых находится небольшое количество фибробластов, внутренний - остеогенный (костеобразующий) - прилежит непосредственно к костной ткани. В нем расположены тонкие веретенообразные «покоящиеся» остеогенные клетки, за счет которых происходят развитие, рост в толщину и регенерация костей после повреждения. В настоящее время остеогенные клетки считаются стволовыми, так как они сохраняют способность к интенсивному делению, самоподдержанию и являются полипотентными, т. е. в процессе дифференцировки из них в зависимости от микроокружения может образоваться несколько типов клеток (остеобласты, хондроциты).
В зависимости от расположения костных пластинок различают плотное (компактное) и губчатое костное вещество (трабекулярная кость) (рис. 9).
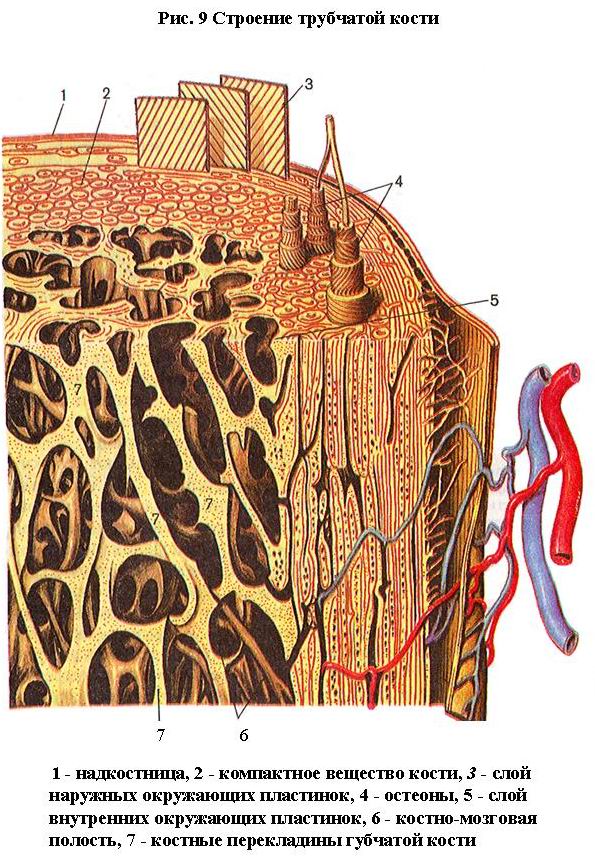
В компактном веществе костные пластинки располагаются в определенном порядке, образуя сложные системы - остеоны. Остеон - структурная единица кости. Он состоит из 5-20 цилиндрических пластинок, вставленных одна в другую. В центре каждого остеона проходит центральный канал (гаверсов) (рис. 10). Диаметр остеона 3-4 мм. Между остеонами залегают интерстициальные (вставочные, промежуточные) пластинки, кнаружи от них находятся наружные окружающие (генеральные) пластинки (рис. 9) . Каналы остеонов сообщаются между собой с помощью коротких поперечных каналов. Снаружи в кость проникают прободающие фолькмановы каналы, в которых проходят сосуды, питающие кость. Между пластинками залегает огромное количество остеоцитов. В центральных каналах проходят кровеносные капилляры. В перикапиллярном пространстве залегают макрофаги, проходят нервные волокна.
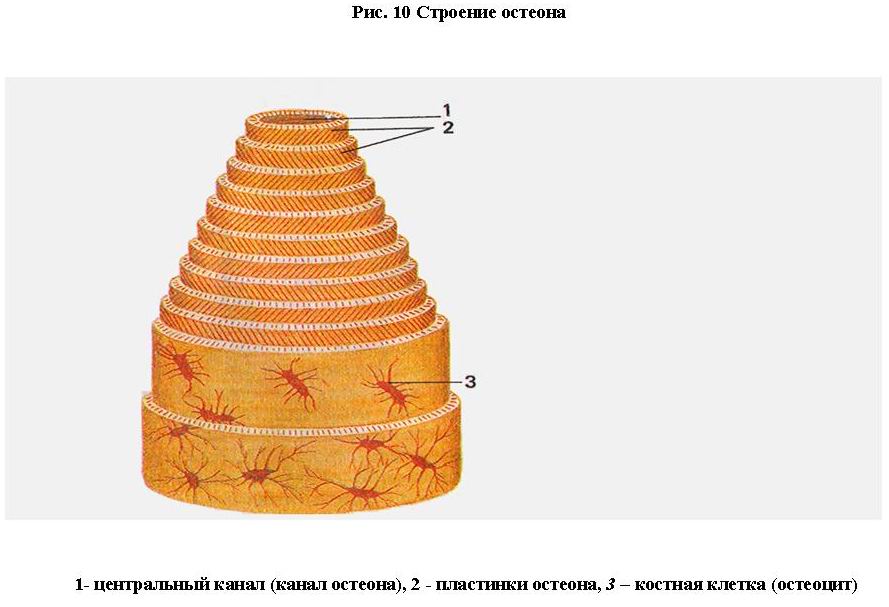
На границе с костномозговой полостью имеется небольшое количество губчатого костного вещества. Губчатое костное вещество состоит из тонких костных перекладин (трабекул), перекрещивающихся между собой и образующих множество ячеек. Перекладины направлены по линиям сжатия и растяжения и образуют сводчатые конструкции (рис. 9) . Такое расположение костных трабекул под углом друг к другу обеспечивает равномерную передачу напряжения, развиваемого мышцей, на кость.
На поверхности каждой кости имеются выпуклости, углубления, борозды, отверстия, шероховатости. Здесь начинаются или прикрепляются мышцы и их сухожилия, фасции, связки, проходят сосуды и нервы. Возвышения, выступающие над поверхностью кости, называют отростками, апофизами. На костях имеются также углубления - ямы, ямки. Поверхности кости ограничены краями. На участках, к которым прилежит нерв или кровеносный сосуд, имеются борозды. В местах прохождения через кость сосуда или нерва образуются канал, каналец, щель, вырезка. На поверхности каждой кости, особенно с внутренней стороны, видны точечные отверстия, уходящие в глубь кости, - питательные отверстия.
Таблица 2.
Классификация костей
Вид кости |
Часть кости |
Трубчатая (длинная, короткая)
Короткая (губчатая) Плоская Смешанная Воздухоносная |
Тело (диафиз), концы (эпифизы), метафизы, отростки, выступы-бугры (апофизы) - Края, углы - Воздухоносная полость |
Кости отличаются друг от друга по форме, при этом их форма и выполняемая функция взаимосвязаны и взаимообусловлены (табл. 2, рис. 11).
У трубчатой кости различают ее удлиненную среднюю часть - тело кости, или диафиз, обычно цилиндрической или близкой к трехгранной формы, и утолщенные концы - эпифизы. На них располагаются суставные, поверхности, покрытые суставным хрящом, служащие для соединения с соседними костями. Участок кости, расположенный между диафизом и эпифизом, называется метафизом. В детском и юношеском возрасте рост костей в длину происходит за счет гиалинового эпифизарного (метаэпифизарного) хряща, который находится между диафизом и эпифизом трубчатой кости.
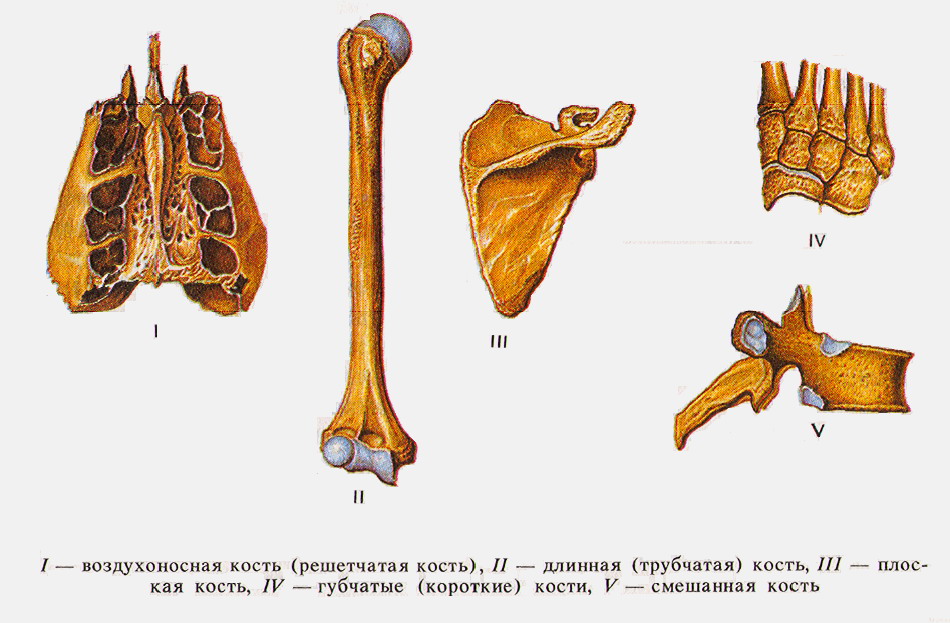
Рис. 11 Виды костей
Среди трубчатых костей выделяются длинные трубчатые кости (плечевая, бедренная, кости предплечья и голени) и короткие (кости пясти, плюсны, фаланги пальцев). Диафизы построены из компактной, эпифизы - из губчатой кости, покрытой тонким слоем компактной.
Губчатые кости, так же как и эпифизы трубчатых костей, состоят из губчатого вещества, покрытого тонким слоем компактного вещества. К этим костям также следует отнести кости, развивающиеся в сухожилиях - сесамовидные (гороховидная, надколенник). Губчатые кости имеют форму неправильного куба или многогранника с закругленными углами и гранями. Такие кости располагаются в местах, где большая нагрузка сочетается с большой подвижностью.
Плоские кости участвуют в образовании полостей, поясов конечностей, выполняют функцию защиты (кости крыши черепа, грудина). К их поверхности прикрепляются мышцы.
Смешанные кости имеют сложную форму. Они состоят из нескольких частей, имеющих различное строение, очертание и происхождение, например позвонки, кости основания черепа.
Воздухоносные кости имеют в своем теле полость, выстланную слизистой оболочкой и заполненную воздухом, например некоторые кости черепа - лобная, клиновидная, решетчатая, верхняя челюсть.
Внутри костей в костномозговых полостях и ячейках губчатого вещества, выстланных эндостом (слоем плоских остеогенных клеток, лежащих на тонкой соединительнотканной пластинке), находится костный мозг. В период внутриутробного развития, а также у новорожденных во всех костных полостях находится красный костный мозг, он выполняет кроветворную и защитную функцию. У взрослого человека красный костный мозг содержится только в ячейках губчатого вещества плоских костей (грудина, крылья подвздошных костей), в губчатых костях и эпифизах трубчатых костей. В диафизах, т. е. в костномозговых полостях, находится желтый костный мозг.
Кость живого человека - динамическая структура, в которой происходят постоянный обмен веществ, анаболические и катаболические процессы, разрушение старых и создание новых костных трабекул и остеонов.
П. Ф. Лесгафт сформулировал ряд общих принципов организации костей:
Костная ткань образуется в местах наибольшего сжатия или натяжения.
Степень развития костей пропорциональна интенсивности деятельности связанных с ними мышц.
Трубчатое и арочное строение кости обеспечивает наибольшую прочность при минимальной затрате костного материала.
Внешняя форма костей зависит от давления на них окружающих тканей и органов, в первую очередь мышц, и меняется при уменьшении или увеличении давления.
Перестройка формы кости происходит под влиянием внешних (для костей) сил.
Кости приспосабливаются к изменяющимся условиям жизнедеятельности организма, под влиянием которых происходит перестройка их макро и микроскопического строения. П. Ф. Лесгафт убедительно показал, что внешняя форма костей меняется под влиянием растяжения и давления, а кости развиваются тем лучше, чем интенсивнее деятельность связанных с ними мышц.
Форма и рельеф костей зависят от характера прикрепления мышц. Так, если мышца прикрепляется к кости с помощью сухожилий, то в этой области формируется бугор, отросток, а если мышца вплетается в надкостницу широким пластом, то формируется углубление (В.А. Долго-Сабуров). В местах прохождения сосудов на костях имеются борозды. Через каналы, щели, канальцы различной величины проходят сосуды и нервы. Поверхность кости испещрена множеством мелких питательных отверстий, через которые внутрь кости проходят сосуды.
На структуру кости оказывает значительное влияние профессия (М. Г. Привес). В зависимости от характера выполняемой работы меняются форма, ширина и длина костей, толщина компактного слоя, размеры костно-мозговой полости и т. д. У лиц, занимающихся тяжелым физическим трудом, позвонки приобретают клиновидную форму, а у балерин или шоферов грузовых машин, постоянно опирающихся на переднюю часть стопы, плюсневые кости утолщены, их костно-мозговые полости сужены. Существенна формообразовательная роль физкультуры и спорта. Все это подтверждает правильность положения П. Ф. Лесгафта о том, что рост и прочность костей определяются интенсивностью деятельности окружающих кость мышц.