

Каротины (провитамины а)
Химическое строение и свойства. Каротины являются изопреноидами и образуются в растениях при фотосинтезе. Известны 3 типа каротинов: α-, β- и γ-каротины.
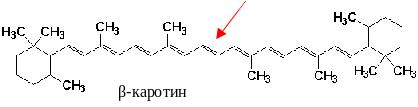
Впервые каротины были выделены из моркови, отсюда и их название (carota – морковь); α- и γ-каротины содержат по одному β-иононовому кольцу, и при их окислительном распаде образуется одна молекула витамина А, в молекуле β-каротина – два β-иононовых кольца, и он обладает большей биологической активностью, поскольку из него образуется две молекулы ретинола:
β -каротин
+ 2 НАД (Ф)H∙H+
2 транс-ретинол
+2 НАД+
-каротин
+ 2 НАД (Ф)H∙H+
2 транс-ретинол
+2 НАД+
Реакция катализируется двумя ферментами: каротиндиоксигеназой, расщепляющей молекулу β-каротина в центральной его части, и редуктазой, с участием коферментов НАДH∙H+ либо НАДФH∙H+ . Одновременный прием с пищей антиоксидантов, препятствующих окислению каротина по периферическим двойным связям, а также витамина В12, повышающего активность каротиндиоксигеназы, увеличивает количество молекул каротина, расщепляющегося по центру, и эффективность синтеза витамина А вследствие этого возрастает в 1,5-2 раза. Усвоению каротина способствуют также флавоноиды.
Главным местом превращения каротина в витамин А является стенка кишечника. Поскольку каротины являются жирорастворимыми соединениями, их усвоение происходит вместе с липидами.
Каротины легко окисляются кислородом воздуха; чувствительны к свету. Процесс их окисления идет аутокаталитически с образованием свободных радикалов. Витамин Е способен предохранять двойные связи в молекуле β-каротина от окисления.
Биохимические функции. Наличие сопряженных двойных связей в молекуле витамина А и каротинов обуславливает их высокую реакционную способность при взаимодействии со свободными радикалами различных типов. В молекуле β-каротина 11 ненасыщенных двойных связей, константа скорости реакции со свободными раликалами у него в 5 раз больше, чем у ретинола. β-каротин способен быть перехватчиком свободных радикалов благодаря стабилизации в его молекуле неспаренного электрона. Таким образом, каротиноиды выполняют роль антиоксидантов, перехватывая синглетный кислород и другие активные формы кислорода (О2˙, ОН˙, Н2О2). β-каротин является наиболее важным гасителем синглетного кислорода. Поскольку облучение и канцерогенез имеют свободнорадикальную природу, β-каротин по праву считается радиозащитным соединением и антиканцерогеном. Он обладат также антимутагенными свойствами. Установлена обратная корреляция межу содержанием в диете β-каротинов и частотой заболеваемости раком. β-каротины синергично взаимодействуют с витамином Е как мембранные антиоксиданты. Они способны депонировать в клетках кислород.
Недостаточность каротинов не описана.
При избыточном потреблении каротинов у человека возможно пожелтение ладоней, подошв стоп и слизистых, однако даже в таких крайних случаях выраженных симптомов ин-токсикации не отмечалось. У животных более чем 100-кратное превышение дозировки β-каротинов увеличивало интенсивность реакций ПОЛ в тканях (прооксидантный эффект), этого не наблюдалось в присутствии витаминов Е и С (защита молекулы каротина от окислительной деструкции).
Суточная потребность в β-каротинах составляет 5 мг.
ВИТАМИН Е
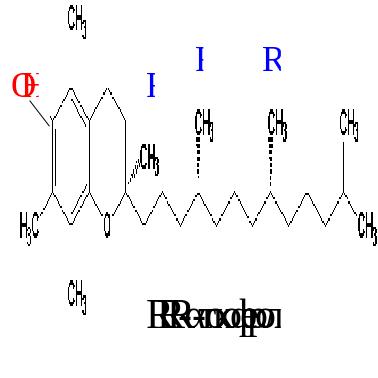
В 1938 г. токоферол, или витамин Е, был синтезирован химическим путем. Однако между природным и синтетическим витамином Е существуют различия, что отражено в их классификации. Натуральные формы токоферола обозначаются как RRR--токоферолы (R обозначает конфигурацию метильной группы) и имеют единственный стереоизомер; синтетические формы называются all-rac- -токоферолы и являются рацемической смесью из 8 стереоизомеров, из которых 7 не найдено в природе. Основной биологически активной формой является RRR-α-токоферол:
Свободная ОН-группа обуславливает свойства витамина Е как антиоксиданта.
Кроме токоферолов к группе витамина Е относятся α, β, γ и δ-токотриенолы – аналоги соответствующих токоферолов, которые отличаются от последних наличием двойных связей в боковой цепи. Токоферолы – прозрачные, светло-желтые, вязкие масла, хорошо растворимые в большинстве органических растворителей. Медленно окисляются на воздухе, разрушаются под действием УФ-лучей
Метаболизм
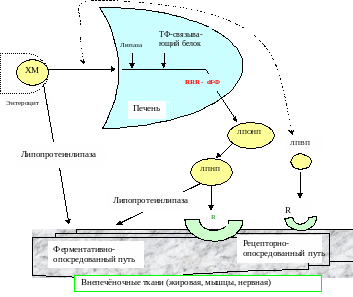
Эстераза+ липаза
Масла
Кровь
Структурная организация фосфолипидов клеточных мембранах способна “узнавать” хиральную форму RRR-α-токоферола, благодаря чему витамин задерживается в мембране, где и выполняет свою функцию
Не всосавшиеся в кишечнике токоферолы выводятся с калом.
Продукты метаболизма витамина – токофериновая кислота и ее водорастворимые глюкурониды – выводятся с мочой.
Биохимические функции.
Витамин Е является универсальным протектором клеточных мембран от окислительного повреждения. Он занимает такое положение в мембране, которое препятствует контакту кислорода с ненасыщенными липидами (образование гидрофобных комплексов). Это защищает биомембраны от их перекисной деструкции. Антиоксидантные свойства токоферола обусловлены также способностью подвижного гидроксила хроманового ядра его молекулы непосредственно взаимодействовать со свободными радикалами кислорода (О2˙, НО˙, НО2˙), свободными радикалами ненасыщенных жирных кислот (RO˙, RO2˙) и пероксидами жирных кислот. Мембраностабилизирующее действие витамина проявляется и в его свойстве предохранять от окисления SH-группы мембранных белков. Его антиоксидантное действие заключается также в способности защищать от окисления двойные связи в молекулах каротина и витамина А. Витамин Е (совместно с аскорбиновой кислотой),способствует включению селена в состав активного центра глутатионпероксидазы, тем самым он активизирует ферментативную антиоксидантную защиту (глутатионпероксидаза обезвреживает гидропероксиды липидов).
/ \ ОО
\ / / / \
/ \ _║¯\ + ОН-ТФ \ / + •O-ТФ
\ / / / / \
/ / / / \
/ \‗/‾ \ \ /
\ / / \‗
/ \ \_
\ / \‗
/ \ \
\ / /
/ \ \
/
Пероксидный радикал ненасы- Ненасыщенная жир- радикал
сыщенной жирной кислоты Токоферол ная кислота токоферола
2. Токоферол является не только антиоксидантом, но и антигипоксантом, что объясняется его способностью стабилизировать митохондриальную мембрану и экономить потребление кислорода клетками. Следует отметить, что из всех клеточных органелл митохондрии наиболее чувствительны к повреждению, так как в них содержится больше всего легко окисляющихся ненасыщенных липидов. Вследствие мембраностабилизирующего эффекта витамина Е в митохондриях увеличивается сопряженность окислительного фосфорилирования, образование АТФ и креатинфосфата. Важно также отметить, что витамин контролирует биосинтез убихинона – компонента дыхательной цепи и главного антиоксиданта митохондрий.
3. Токоферол контролирует синтез нуклеиновых кислот (на уровне транскрипции), а также гема, микросомных цитохромов и других гем-содержащих белков.
4. Витамин Е обладает способностью угнетать активность фосфолипазы А2 лизосом, разрушающей фосфолипиды мембран. Повреждение мембран лизосом приводит к выходу в цитозоль протеолитических ферментов, которые и повреждают клетку.
5. Витамин Е является эффективным иммуномодулятором, способствующим укреплению иммунозащитных сил организма.
Гиповитаминоз. Недостаточность токоферола – весьма распространенное явление, особенно у людей, проживающих на загрязненных радионуклидами территориях, а также подвергающихся воздействию химических токсикантов. Глубокий гиповитаминоз встречается редко – преимущественно у недоношенных детей (проявляется гемолитической анемией).
При Е-витаминной недостаточности наблюдается частичный гемолиз эритроцитов, в них снижается активность ферментов антиоксидантной защиты. Повышение проницаемости мембран всех клеток и субклеточных структур, накопление в них продуктов ПОЛ – главное проявление гиповитаминоза. Именно этим обстоятельством объясняется разнообразие симптомов недостаточности токоферола. – от мышечной дистрофии и бесплодия вплоть до некроза печени и размягчения участков мозга, особенно мозжечка. Увеличение активности выходящих из поврежденных тканей ферментов в сыворотке крови (креатинфосфокиназы, аланинаминотрансферазы и др.) и увеличение содержания в ней продуктов ПОЛ наблюдается уже на ранних стадиях Е-гиповитаминоза.
Дефицит витамина Е в организме сопровождается снижением содержания иммуноглобулинов E. После его введения нормализуется численность Т- и В-лимфоцитов в периферической крови и восстанавливается функциональная активность Т-клеток.
Описаны врожденные нарушения обмена витамина Е, проявляющиеся анемией и мышечной дистрофией.
Оценка обеспеченности организма токоферолом. Основным методом является определение концентрации витамина Е в крови. Однако при гипер-β-липопротеинемиях (в том числе и возрастных) содержание токоферола в крови не отражает уровень обеспеченности им тканей организма (ЛПНП – одна из транспортных форм витамина).
Наиболее доступны клинической лаборатории функциональные тесты, позволяющие оценить глубину Е-витаминной недостаточности. Основными являются тесты на перокидную или гемолитическую устойчивость эритроцитов, определение суточной экскреции креатина (мышцы при Е-гиповитаминозе плохо «удерживают» креатин) и содержание продуктов ПОЛ в крови и эритроцитах.
Гипервитаминоз. Витамин не токсичен при значительных (10-20-кратных к суточной потребности) и длительных превышениях его дозировки, что обусловлено ограничением способности специфических токоферолсвязывающих белков печени включать витамин в состав ЛПОНП. Его избыток выводится из организма с желчью. В некоторых случаях длительный прием мегадоз токоферола (более 1г/сут) может привести к гипертриглицеридемии и повышению кровяного давления.
Суточная потребность – 10 мг. Однако токоферол быстро расходуется в организме, особенно в условиях стимулированного ПОЛ, что имеет место при многих заболеваниях. Окислительной деструкции витамина Е препятствует витамин С. Это обусловлено присутствием в молекуле последнего енольной группы - донора водорода, который способен «гасить» свободный радикал токоферола. Тем самым витамин С экономит фонд витамина Е. С целью усиления антиоксидантного эффекта токоферола его следует назначать с аскорбиновой кислотой.
ВИТАМИН Д (КАЛЬЦИФЕРОЛ), АНТИРАХИТИЧЕСКИЙ.
Эргостерин, витамин Д1.
витамин Д2, эргокальциферол (кальциферол означает несущий кальций). Образуется при воздействии на витамин Д1 УФ-лучей.
Витамин Д3 , Холекальциферол
витамеры эргостерина (Д4 -7), синтезируются в растениях при УФ-облучении
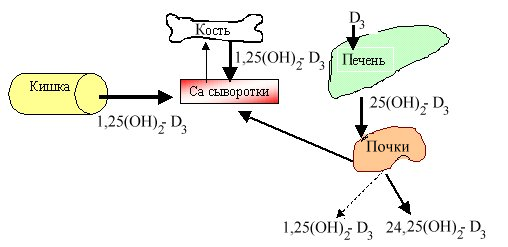
Метаболизм
Холестерол
7-дегидрохолестерол
Кожа
УФ-лучи
Холекальциферол
Печень
25(ОН)-Д3,
25-гидроксихолекальциферол
Гидроксилаза-1
Почки
Гидроксилаза-1
Микросомальная система
Митохондриальный фермент
1,25(ОН)2-Д3,
1,25-дигидроксихолекальциферол, или кальцитриол
Паратгормон.
В случае когда уровень Са в крови снижен
24,25(ОН)2-Д3,
24,25-дигидроксихолекальциферол
Если уровень Са адекватен физиологической потребности организма,
Вит С
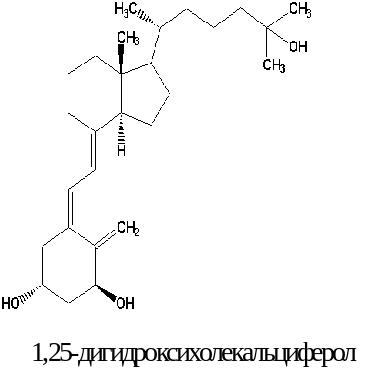
Биохимические функции. Витамин Д3 можно рассматривать как прогормон, так как он превращается в 1,25(ОН)2-Д3, действующий аналогично стероидным гормонам. Так, проникая в клетки-мишени, он связывается с белковыми рецепторами, которые мигрируют в ядро клетки. В энтероцитах этот гормон-рецепторный комплекс стимулирует транскрипцию иРНК, несущую информацию на синтез белка-переносчика ионов кальция. Вероятно, витамин отвечает также за синтез Са2+-АТФ-азы в разных клетках.
В кишечнике всасывание кальция осуществляется как путем облегченной диффузии (с участием кальцийсвязывающего белка), так и путем активного транспорта (с помощью Са2+-АТФ-азы). Одновременно ускоряется и всасывание фосфора.
В костной ткани 1,25(ОН)2-Д3 стимулирует процесс деминерализации (синергично с паратирином).
В почках активация витамином 1,25(ОН)2-Д3 кальциевой АТФ-азы мембран почечных канальцев приводит к увеличению реабсорбции ионов кальция; возрастает и реабсорбция фосфатов.
Кальцитриол принимает участие в регуляции роста и дифференцировке клеток костного мозга. Он обладает антиоксидантным и антиканцерогенным действием.
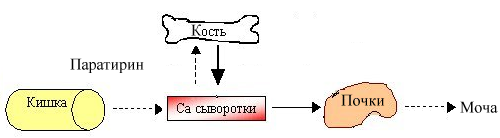
Гиповитаминоз. Недостаток витамина Д у детей приводит к заболеванию рахитом. Основные проявления этого заболевания сводятся к симптоматике недостаточности кальция. Прежде всего страдает остеогенез: отмечается деформация скелета конечностей (искривление их в результате размягчения – остеомаляции), черепа (позднее заращение родничков), грудной клетки (появление своеобразных «четок» на костно-хрящевой границе ребер), задерживается прорезывание зубов. Развивается гипотония мышц (увеличенный живот), возрастает нервно-мышечная возбудимость (у младенца выявляется симптом облысения затылочка из-за частого вращения головкой), возможно появление судорог. У взрослого недостаточность кальция в организме приводит к кариесу и остеомаляции (размягчение кости); у пожилых – к развитию остеопороза (снижение плотности костной ткани вследствие нарушения остеосинтеза). Разрушение неорганического матрикса объясняется усиленным «вымыванием» кальция из костной ткани и нарушением реабсорбции кальция в почечных канальцах при дефиците витамина Д:
На схеме показано угнетение (пунктирная стрелка) всасывания, снижение поступления кальция в кость и уменьшение экскреции кальция при недостатке витамина Д. Одновременно в ответ на гипокальциемию секретируется паратирин и увеличивается (сплошная стрелка) поступление кальция из кости в кровяное русло (вторичный гиперпаратиреоидизм).
Врожденные нарушения обмена витамина Д
Семейный гипофосфатемический витамин- Д-резистентный рахит. Клиническое течение этой формы рахита весьма вариабельно: от некоторой задержки роста ребенка до развития симптомов тяжелого рахита. Изменения скелета обычно проявляются на 1-2 году жизни, типична «утиная» походка, однако мышечная гипотония отсутствует. Задержка роста и утолщение костей при этом заболевании приводят к формированию приземистого, псевдоатлетического телосложения.
Основным биохимическим симптомом является гипофосфатемия в результате снижения реабсорбции неорганического фосфата в почечных канальцах. Фосфор теряется с мочой. Нарушается также всасывание фосфора (нередко и кальция) в тонком кишечнике. Избирательный дефект реабсорбции фосфата в почечных канальцах связан не с нарушением образования гормонально-активной формы витамина Д, а с генетически обусловленным повреждением структуры белков, участвующих в этом процессе.
Лечение: назначение высоких доз витамина Д и фосфатов.
Врожденный псевдодефицитный витамин-Д-зависимый рахит..Клиническая картина подобна проявлениям «классического» рахита. В крови снижено содержание кальция, иногда – фосфора, резко повышена активность щелочной фосфатазы в крови (повышение активности этого фермента является диагностическим показателем). Характерны гипераминоацидурия и вторичный гиперпаратиреоидизм.
Главным метаболическим дефектом является нарушение образования в почках 1,25-D3 (кальцитриола), вследствие чего нарушается всасывание кальция в тонком кишечнике. Развивающаяся гипокальциемия ведет к вторичному гиперпаратиреоидизму.
Лечение: парэнтеральное введение 1,25(ОН)2-Д3, либо постоянное назначение высоких (1000-кратно превышающих физиологические) доз холекальциферола.
Гипервитаминоз. Избыточный прием витамина Д приводит к интоксикации и сопровождается выраженной деминерализации костей – вплоть до их переломов. Содержание кальция в крови повышается. Это сопровождается кальцификацией мягких тканей, особенно склонны к этому процессу почки (образование камней, развитие почечной недостаточности).
Повышение уровня кальция (и фосфора) в крови объясняется следующим образом : 1) резорбцией костной ткани (сплошная стрелка); 2)увеличением интенсивности всасывания кальция и фосфора в кишечнике; 3)увеличением их реабсорбции в почках (т.е. угнетение экскреции с мочой – пунктир).
В нормальных условиях повышение содержания кальция в крови будет приводить к образованию неактивного 24,25(ОН)2-Д3, который не вызывает резорбцию («рассасывание») кости, однако при гипервитаминозе Д этот механизм становится неэффективным.
Интересно, что пигментация кожи (загар) является защитным фактором, предохраняющим от избыточного образования витамина Д при УФ-облучении кожи. Однако у светлокожих жителей северных стран несмотря на недостаток солнечной инсоляции, витамин Д-дефицитные состояния, как правило, не развиваются, так как их диета включает рыбий жир.
Оценка обеспеченности организма витамином Д. Обеспеченность организма витамином Д оценивается на основании определения: 1)активных форм витамина Д в крови и тканях методом радиоконкурентного анализа; 2) содержания кальция, фосфора и активности щелочной фосфатазы в сыворотке крови; 3)уровня экскреции с мочой фосфатов. Применяются также нагрузочные пробы с приемом фиксированных доз кальция при парэнтеральном введении с последующим определением содержания кальция в крови и моче.
Суточная потребность для детей колеблется от 10 до 25 мкг (500-1000 МЕ), у взрослых она ниже
ВИТАМИН К (НАФТОХИНОНЫ), АНТИГЕМОРРАГИЧЕСКИЙ.
Витамин К – это две группы хинонов с боковыми изопреноидными цепями:
витамин К1 (филлохиноны) в растениях
витамин К2 (менахиноны). имеется также и у животных.
Они различаются строением и количеством изопреновых единиц в боковой цепи.
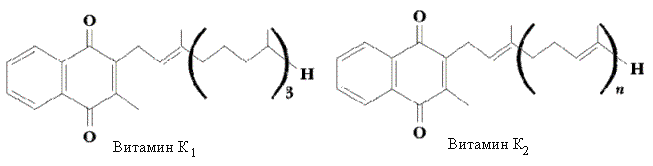
Оба витамина не растворяются в воде, но хорошо растворимы в органических растворителях; они разрушаются при нагревании в щелочной среде и на свету.
Синтезированы аналоги витамина К – менадион (витамин К3, лишенный боковой цепи) и его водорастворимый аналог викасол, последний широко применяется в медицинской практике.
Метаболизм. Витамин К всасывается аналогично всем жирорастворимым витаминам, т.е. включается вначале в состав мицелл, а затем – хиломикронов. В плазме крови он связывается с альбуминами. Накапливается в печени, селезенке и сердце. В тканях образуется активная форма витамина – менахинон-4 (содержит 4 изопреноидные единицы). Конечные продукты обмена витамина выделяются с мочой.
Биохимические функции. Единственная известная биологическая роль витамина К заключается в том, что он является коферментом γ-глутамилкарбоксилазы, карбоксилирующей глутаминовую кислоту с образованием γ-карбоксиглутаминовой кислоты.
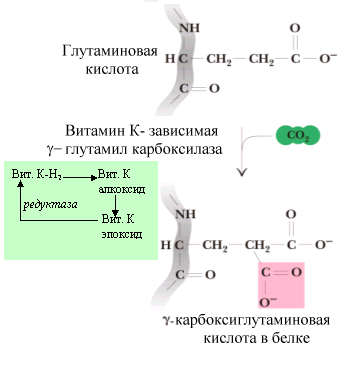
Фермент удалось очистить до гомогенного состояния (белковая цепь содержит 758 аминокислот с пробладанием гидрофобных) и определить его структурный ген (он включает 15 экзонов).
При взаимодействии восстановленной формы витамина К-Н2 с γ-глутамилкарбоксилазой в присутствии кислорода образуется сильное основание (алкоксид), способное отнять от -С-атома глутаминовой кислоты водород, на место которого присоединяется СО2 – при этом образуется γ-карбоксиглутаминовая кислота. В ходе реакции появляются короткоживущие и высокотоксичные промежуточные соединения (свободные радикалы витамина К), которые превращаются в нетоксичный эпоксид витамина К в присутствие глутамата. Эпоксид витамина К снова восстанавливается в витамин К-Н2 с помощью ферментов редуктаз. Следовательно, при дефиците глутаминовой кислоты в клетке затрудняется обезвреживание токсичных свободных радикалов витамина К.
-карбоксиглутаминовая кислота является Са++-связывающей аминокислотой, которая необходима для функционирования кальцийсвязывающих белков. К таковым относятся:
факторы свертывающей системы крови: IX, VII, X и протромбин;
регуляторные белки (протеин С и протеин S), нуждающиеся в -карбоксиглутаминовой кислоте для Са-индуцированного взаимодействия с поверхностью клеточной мембраны;
белки минерализации костной ткани (костный -карбоксиглутаминовый протеин и другие). Поскольку при дефекте синтеза костного -карбоксиглутаминового белка кальцифицируются артерии и хрящи, возможно, что его функцией является также контроль за внекостной кальцификацией;
витамин К-зависимый белок Gas 6, активирующий рост гладкомышечных клеток; витамин К-зависимый сократительный белок хвостика сперматозоида;
некоторые нейротоксины (например, содержащиеся в яде улитки).
Общей особенностью всех витамин-К-зависимых белков является формирование белковой сеточки, образованной γ-карбоксиглутаминовой кислотой, связанной с кальцием. Такая сеточка впервые была описана для протромбина. Протромбин в присутствие Са++ связывается с биомембраной, что является необходимым условием для реализации процесса свертывания крови.
Гиповитаминоз. Признаком недостаточности витамина К является повышенная кровоточивость, особенно при травмах. У взрослого человека гиповитаминоз К встречается довольно редко, так как этот витамин имеется во многих пищевых продуктах; кроме того, он синтезируется микрофлорой кишечника. У новорожденных недостаточность витамина К (геморрагическая болезнь новорожденнных) – явление нередкое из-за его низкого содержания в материнском молоке и недостаточно развитой микрофлоры кишечника. Гиповитаминоз жирорастворимых витаминов может развиваться при нарушении выделения желчи, панкреатитах, дисбактериозах кишечника и некоторых других заболеваниях желудочно-кишечного тракта.
В связи с участием витамина К в остеосинтезе можно полагать, что недостаток витамина К играет роль в развитии остеопороза, т.е. разрежения и истончения структуры кости.
Врожденные нарушения обмена витамина К
Врожденный дефицит факторов II (протромбина), VII, IX и X. В основе заболевания лежит дефект синтеза контролируемых витамином К белков свертывающей системы крови. Резко увеличивается протромбиновое время.
Врожденная резистентность к антагонистам витамина К. В основе заболевания лежит генетическая мутация структуры белкового рецептора, связывающего витамин К и его антагонистов. Вследствие этого возникает повышенная потребность в витамине и увеличивается резистентность организма к антикоагулянтам.
Гипервитаминоз К не описан.
Оценка обеспеченности организма витамином К. Определение содержания витамина К осуществляется физико-химическими, радиоизотопными и биологическими методами (в эксперименте на животных).
Суточная потребность – приблизительно 0,1 мг/сутки.