

Билет 10
Витамин PP существует в двух формах - никотиновой кислоты и никотиномида. Активное воздействие витамина PP на обменные процессы обусловлено его вхождением в состав ниацинамидадениндинуклеотида (НАД) и ниацинамидадениндинуклеотида фосфата (НАДФ), являющихся кофакторами ряда ферментов. Никотиновая кислота в организме входит в состав NAD и NADP, выполняющих функции коферментов различных дегидрогеназ (см. раздел 2). Синтез NAD в организме протекает в 2 этапа:
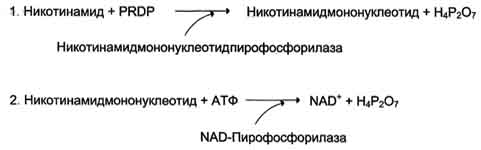
NADP образуется из NAD путём фосфорилирования под действием цитоплазматической NAD-киназы.
NAD+ + АТФ → NADP+ + АДФ
В частности, ниацинамид входит в состав кодегидраз, являющихся переносчиками водорода к флавопротеиновым ферментам, и тем самым регулирует окислительно-восстановительные процессы в организме. Он участвует более чем в полусотне реакций, в ходе которых сахар и жир превращаются в энергию. Он также необходим для обмена аминокислот и участвует в превращении жиров в вещества, именуемые эйкозаноидами, - гормоноподобные агенты, управляющие метаболическими путями нашего организма. Ниацин - витамин, не знающий равных в контроле холестерина. Ниацин борется с такими факторами риска сердечно-сосудистых заболеваний
Ниацин это белый кристаллический порошок без запаха, слабокислого вкуса. Трудно растворим в холодной воде (1:70), лучше в горячей (1:15), мало растворим в этаноле, очень мало — в эфире. Суточная потребность взрослого человека 15—20 мг.
Источники. Витамин РР широко распространён в растительных продуктах, высоко его содержание в рисовых и пшеничных отрубях, дрожжах, много витамина в печени и почках крупного рогатого скота и свиней. Витамин РР может образовываться из триптофана (из 60 молекул триптофана может образоваться 1 молекула никотинамида), что снижает потребность в витамине РР при увеличении количества триптофана в пище.
Недостаточность витамина РР приводит к заболеванию "пеллагра", для которого характерны 3 основных признака: дерматит, диарея, деменция ("три Д"), Пеллагра проявляется в виде симметричного дерматита на участках кожи, доступных действию солнечных лучей, расстройств ЖКТ (диарея) и воспалительных поражений слизистых оболочек рта и языка. В далеко зашедших случаях пеллагры наблюдают расстройства ЦНС (деменция): потеря памяти, галлюцинации и бред.
2. Гликоген служит в животном организме резервом углеводов, из которого по мере метаболической потребности могут высвобождаться глюкозофосфат или глюкоза. Хранение в организме собственно глюкозы неприемлемо из-за ее высокой растворимости: высокие концентрации глюкозы создают в клетке высоко гипертоническую среду, что приводит к притоку воды. Напротив, нерастворимый гликоген осмотически почти неактивен.
Гликоген
животных, как и амилопектин растений,
представляет собой разветвленный
гомополимер глюкозы, в котором остатки
глюкозы соединены α(1→4)-гликозидной
связью. Связи в точках ветвления находятся
в положении
α(1→6)
примерно каждого 10-го остатка. Таким
образом, возникает древовидная структура
с молекулярной массой
>1ּ107
Да (до 50 000 остатков), в которой имеется
только одна свободная аномерная
ОН-группа, т. е. только один восстанавливающий
конец.
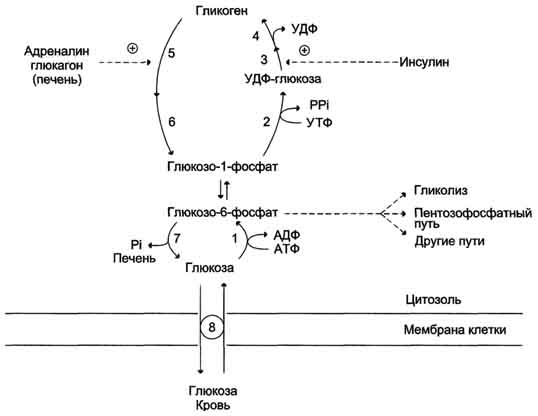
Синтез и распад гликогена. 1 - гексокиназа или глюкокиназа (печень); 2 - УДФ-глюкопирофосфорилаза; 3 - гликогенсинтаза; 4 - амило-1,4 → 1,6-глюкозилтрансфераза (фермент ветвления); 5 - гликогенфосфорилаза; 6 - "деветвящий" фермент; 7 - глюкозо-6-фосфатаза (печень); 8 - транспортные системы ГЛЮТ.
Гликоген печени никогда не расщепляется полностью. Как правило, укорачиваются или удлиняются (при высоком содержании глюкозы) только невосстанавливающие концы древовидной структуры. Удлинение цепи катализируется гликоген-синтазой [2]. Так как образование гликозидных связей между сахарами является эндоэргической реакцией, вначале в реакции глюкозо-1-фосфата с уридинтрифосфатом [УТФ (UTP)] образуется активированный предшественник — УДФ-глюкоза (UDP-глюкоза) ([1]). После этого остаток глюкозы легко переносится с этого промежуточного соединения на гликоген. Когда растущая цепь достигает определенной длины (>11 остатков), специальный фермент ветвления гликогена (1,4→1,6-трансгликозидаза ) [3] катализирует перенос концевого олигосахарида, состоящего из 6-7 остатков, на 6-ОН остаток глюкозы той же или другой цепи гликогена с образованием точки ветвления [α(1→6)-связи] Дальнейшее удлинение этого фрагмента осуществляется гликоген-синтазой, образующей α(1→4)-связи.
Разветвленная структура гликогена облегчает быстрое освобождение углеводных остатков. Наиболее важным ферментом деградации гликогена является гликоген-фосфорилаза [4], отщепляющая от невосстанавливающего конца цепи остатки глюкозы в виде глюкозо-1-фосфата. Чем больше таких концов, тем больше молекул фосфорилазы могут действовать одновременно. Образование глюкозо-1-фосфата вместо глюкозы имеет то преимущество, что для включения освобожденных остатков глюкозы в гликолиз или ГМП не требуется АТФ.Благодаря структуре гликоген-фосфорилазы, процесс последовательного отщепления останавливается за 4 остатка глюкозы от точки разветвления. Точки ветвления удаляются двумя другими ферментами [5 и 6]. Вначале трисахарид боковой цепи переносится [5] к невосстанавливающему концу главной цепи. Затем 1,6-гликозидаза [6] отщепляет остающийся единичный остаток глюкозы в точке ветвления в виде свободной глюкозы, после чего неразветвленная цепь, может вновь расщепляться фосфорилазой.
В организме человека может содержаться до 450 г гликогена, треть из которого накапливается в печени, а остальное — главным образом в мышцах. Содержание гликогена в других органах незначительно. Гликоген печени служит прежде всего для поддержания уровня глюкозы в крови в фазе пострезорбции. Поэтому содержание гликогена в печени варьирует в широких пределах. При длительном голодании оно падает почти до нуля, после чего начинается снабжение организма глюкозой с помощью глюконеогенеза. Гликоген мышц служит резервом энергии и не участвует в регуляции уровня глюкозы в крови. В мышцах отсутствует глюкозо-6-фосфатаза, поэтому гликоген мышц не может быть источником глюкозы в крови. По этой причине колебания содержания гликогена в мышцах меньше, чем в печени.
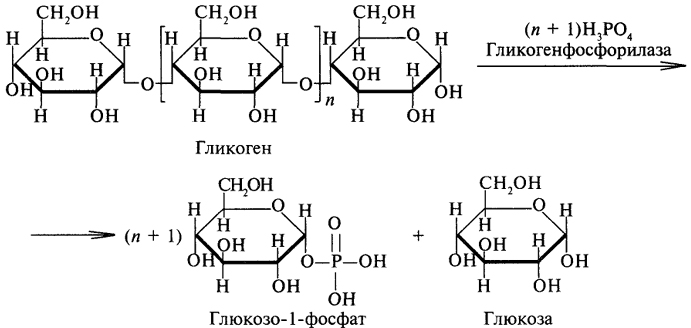
Фосфоролиз является основным путем распада гликогена, его катализирует фермент гликогенфосфорилаза, относящийся к классу трансфераз. Гликогенфосфорилаза отщепляет остатки глюкозы с нередуцирующего конца гликогена и переносит их на молекулу фосфорной кислоты с образованием глюкозо-1-фосфата:
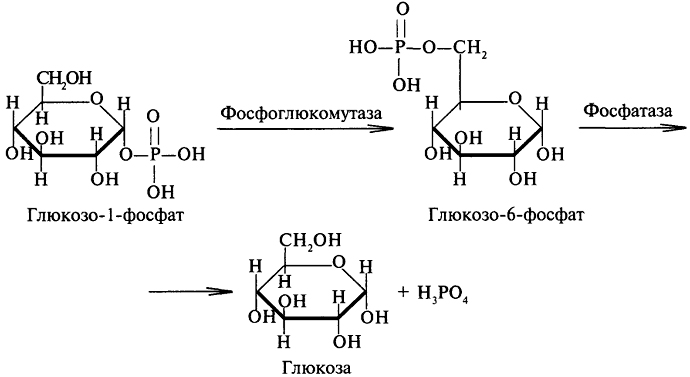
Глюкозо-1-фосфат быстро изомеризуется, превращаясь в глюкозо-6-фосфат, который в печени гидролизуется фосфатазами до глюкозы и фосфорной кислоты:
Процесс фосфоролиза гликогена тонко регулируется. Регуляция активности гликогенфосфорилазы носит каскадный характер, в котором можно выделить несколько видов регуляции ферментативной активности:
гормональная (глюкагон в печени, адреналин в мышцах);2) аллостерическая;3) протеинкиназные реакции (в данном случае - фосфорилирование бокового радикала серина в гликогенфосфорилазе).
Активность мышечной фосфорилазы увеличивается при определенной концентрации АМФ и ацетилхолина, а также в присутствии катионов кальция и натрия.Снижение скорости фосфоролиза происходит при уменьшении запасов гликогена и фосфорной кислоты, а также при увеличении концентрации глюкозо-6-фосфата. Механизмы, снижающие скорость фосфоролиза гликогена, предохраняют организм от больших трат углеводных запасов (гликогена), которые могли бы привести к недостатку глюкозы, необходимой для работы головного мозга и сердечной мышцы.
3. Клеточная мембрана - это неотъемлемый компонент любой клетки. Ее роль в первую очередь состоит в том, чтобы отграничить внутреннее пространство клетки от внешней среды, а у эукариот, кроме того, разделить внутреннюю часть клетки на функционально значимые отсеки: ядро и митохонодрии. В мембранах содержатся липиды трех классов: фосфолипиды, холестерин и гликолипиды. Наиболее важная группа, фосфолипиды, включает фосфатидилхолин (лецитин), фосфатидилэтаноламин, фосфатидилсерин, фосфатидилинозит и сфингомиелин. Холестерин присутствует во внутриклеточных мембранах животных клеток (за исключением внутренней мембраны митохондрий). Гликолипиды входят в состав многих мембран (например, во внешний слой плазматических мембран). В состав гликолипидов входят углеводные функциональные группы которые ориентируются в водную фазу.
Липиды мембран представляют собой амфифильные молекулы с полярной гидрофильной головкой (голубого цвета) и неполярным липофильным хвостом (желтого цвета). В водной среде они агрегируют за счет гидрофобных взаимодействий и вандерваальсовых сил.
Даниелли в связи с необходимостью объяснить явное расхождение между поверхностным натяжением на границах раздела масло/вода и мембра-на/вода. Была высказана гипотеза, что мембрана состоит из двойного липидного слоя, и предположено, что белок располагается на ее поверхности – модель Даниели – Дэвсона, или модель «сэндвича»). Это была очень удачная модель, и в течение последующих 30 лет многочисленные экспериментальные данные, особенно полученные с помощью дифракции рентгеновских лучей и электронной микроскопии, полностью подтвердили ее адекватность. Основными компонентами биологической мембраны являются липид и белок, вопрос о взаимном расположении этих компонентов в мембране стал предметом многочисленных дискуссий, так как обнаружилось, что мембраны выполняют разнообразные функции.
ЖИДКОСТНО-МОЗАИЧНАЯ МОДЕЛЬ МЕМБРАН КЛЕТКИ
Данная модель основана на предшествующих моделях структурно-функциональной организации мембран клетки. Живаямембрана представляет собой двумерный раствор глобулярных интегральных белков, диспергированных в жидкомфосфолипидном матриксе. Экспериментальные подтверждения данного предположения были получены при искусственно вызванном слиянии двух разных родительских клеток. При образовании плазматической мембраны гибридной клетки происходит быстрое стохастическое перемещение с систематическим упорядочением видоспецифичных белков и фосфолипидов. Такие перемещения в плоскости мембраны были названы латеральной подвижностью (диффузией) компонентов мембран.