

2.4.1. Структурные элементы нуклеиновых кислот
Помимо их роли как структурных элементов нуклеиновых кислот нуклеотиды и их производные представляют и самостоятельный биологический интерес. Все нуклеотиды построены из трех компонентов: фосфорной кислоты, альдопентоз рибозы или дезоксирибозы и азотистого основания, обычно производного пурина или пиримидина.

Эти три компонента образуют два типа нуклеотидов, отличающихся природой остатка альдопентозы. Рибонуклеиновые кислоты (РНК) представляют собой полинуклеотиды, содержащие остаток рибозы, а полимерные цепи дезоксирибо-нуклеиновых кислот (ДНК) построены с участием моносахарида дезоксирибозы. На рис. 2.15 изображены также химические формулы азотистых оснований, входящих в состав нуклеотидов ДНК. Три из этих оснований, аденин (А), гуанин (G) и цитозин (С), типичны как для ДНК, так и для РНК. Напротив, тимин (Т) входит в состав только ДНК, а родственное пиримидиновое основание урацил (U) специфично для РНК. Оба типа нуклеотидов представляют собой сильные кислоты, что обусловлено наличием остатка фосфорной кислоты.
Отщепление фосфатной группы от 5'-углеродного атома нуклеотида приводит к соответствующему нуклеозиду. Как показано в табл. 2.10, названия нуклеозидов и нуклеотидов явля ются производными от названий соответствующих азотистых оснований. Следует отметить, однако, что для нуклеотидов применяется и другая номенклатура. Например, аденилат можно назвать также аденозин-5'-монофосфатом. Номенклатура последнего типа применяется обычно для производных нуклеози-дов, у которых гидроксильная группа при С-5' этерифицирована дифосфатной или трифосфатной группировкой, например адено-зин-5'-трифосфат (АТР).
С биологической точки зрения особенно
важен нуклеозид аденозин,
построенный
из остатков рибозы и аденина. На
биологической точки зрения особенно
важен нуклеозид аденозин,
построенный
из остатков рибозы и аденина. На
рис. 2.16 изображено строение аденозин-5'-монофосфата (AMP) и ряда его важных производных. К AMP могут быть присоединены еще один или два остатка фосфорной кислоты, в результате чего образуются ADP (аденозин-5'-дифосфат) и АТР соответственно. При гидролизе соединяющих фосфатные группы фосфодиэфирных связей высвобождается большое количество энергии. Например, превращение АТР в ADP и фосфат при 3°С и рН 7 (напомним, что рН=— IgaH, где an — концентрация ионов Н+ в растворе в моль/л) сопровождается изменением стандартной энергии Гиббса, равным —7,3 ккал/моль.
Мы привыкли оценивать энергетический эффект реакций прежде всего в единицах тепловой энергии, т. е. теплоты, однако клетка представляет собой, в сущности, изотермическую систему, в которой, как правило, реализуются химические пути трансформации энергии. Позднее, мы значительно подробнее рассмотрим АТР в качестве основного переносчика химической энергии во всех клетках без исключения. По сути дела, АТР является аккумулятором энергии, получаемой из питательных веществ или солнечного света, которая затем расходуется в биосинтезе полимеров, транспорте веществ через мембраны и движении клеток. Дифосфаты и трифосфаты других нуклеотидов также могут выполнять аналогичные функции

РИС 2 16 Фосфаты аденозина. AMP, ADP и АТР участвуют в процессах переносаэнертин в метке, а циклический AMP выполняет регулярные функ-ции.
в химии клетки, но основными переносчиками энергии служат все же аденозинфосфаты.
Циклическая форма AMP, содержащая внутримолекулярный цикл с участием фосфатной группы
(рис. 2.16), выполняет функции регулятора множества клеточных реакций, включая реакции образования полисахаридов и резервных полимеров (жиров).
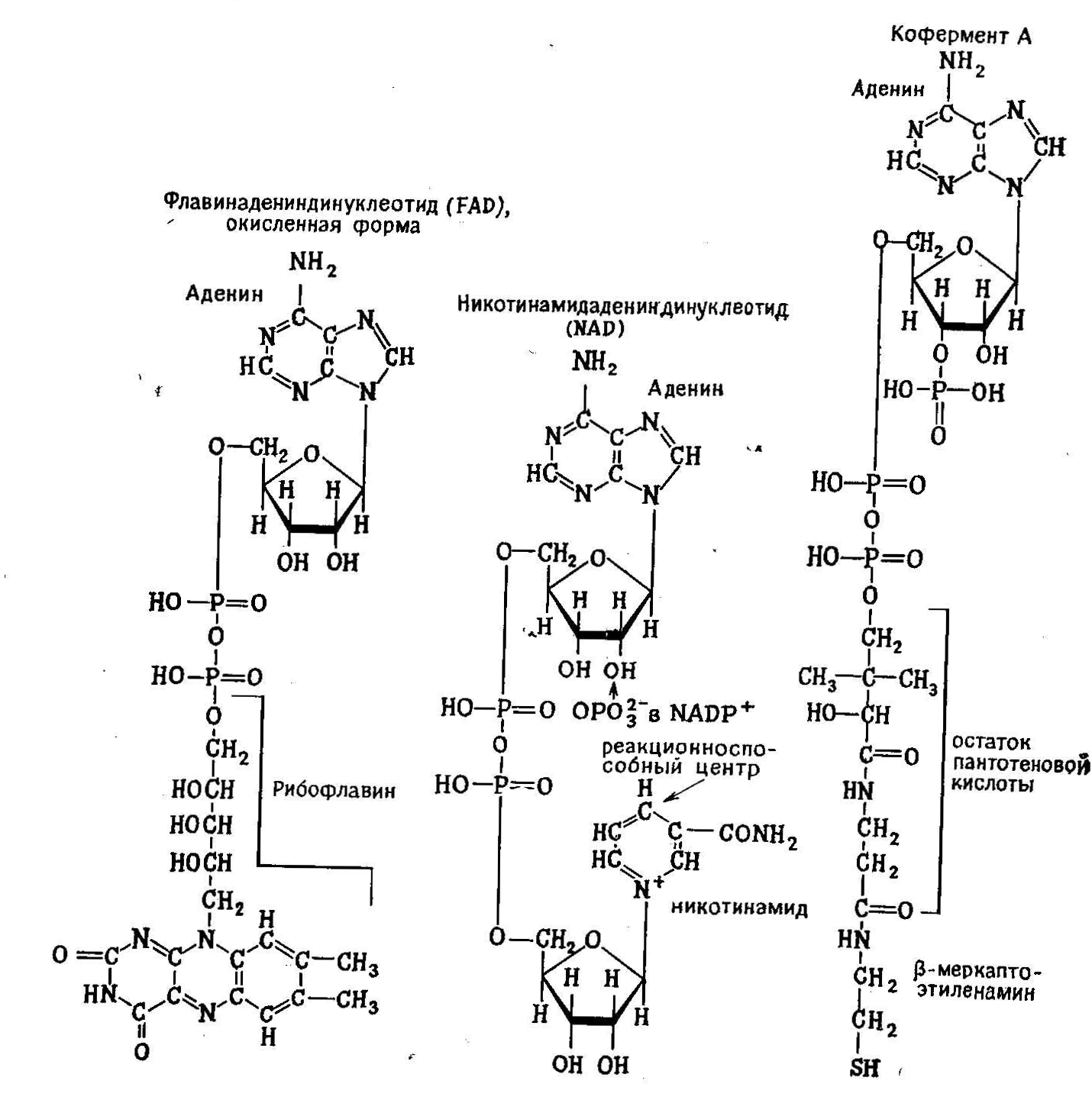
РИС. 2.17. Три важных кофермента, являющиеся производными нуклеотидов.
Недостаток циклического AMP в тканях связывают с одним из видов рака, т. е. состояния относительно неконтролируемого роста клеток.
Аденозинмонофосфат не только является структурным элементом нуклеиновых кислот, но и служит основой для построения ряда коферментов, химические формулы которых изображены на рис. 2.9. Кинетика ферментативных реакций рассматривается в следующей главе; здесь же достаточно отметить, что коферментами мы называем органические соединения, необходимые для активации некоторых ферментов, т. е. для их перевода в ту форму, в которой они способны выполнять каталитические функции. Поскольку практически все реакции клетке катализируются ферментами, изменение концентраций 1оферментов представляет собой удобный способ регуляции активности соответствующих ферментов в клетке и таким путем изменения скорости некоторых внутриклеточных процессов.