

Скаты — batomorpha
Тело уплощено в спинно-боковом направлении. У многих форм хвостовой стебель развит слабо, а лопасть хвостового плавника может редуцироваться. Грудные плавники сильно увеличены, с широкими основаниями. 5 пар жаберных щелей расположены на брюшной стороне. Зубы обычно имеют вид невысоких призм, плотно прилегающих друг к другу и образующих мощную «терку». Сюда относится пять отрядов. Например,
Отряд Пилорылообразные скаты, или Пилы-рыбы, — Pristiformes
Отряд состоит из одного рода и 7 видов пил-рыб. Достигают 5—6 м длины; тело слабо уплощено, грудные плавники относительно невелики. Жаберные щели расположены на нижней поверхности головы.
Подкласс Цельноголовые, отряд Химерообразные — Chimaeriformes
Вальковатое тело несколько сжато с боков и заметно утончается к хвосту. У высокого первого спинного плавника имеется острый шип; у некоторых видов у его основания лежит ядовитая железа. Хвост гетероцеркальный или продолжается в виде длинного утончающегося бича. Морские, преимущественно глубоководные формы. Распространены в умеренных и теплых водах Мирового океана. Лишь немногие виды образуют промысловые скопления. Держатся у дна; мощными зубами легко разгрызают панцири ракообразных и иглокожих, крепкие раковины моллюсков. Плавают за счет волнообразных движений грудных плавников и боковых изгибов хвоста. Длина от 60 см до 1,5—2 м. Размножение растянутое. Одновременно самка откладывает лишь одно-два крупных яйца, каждое из которых заключено в удлиненную овальную роговую капсулу (длиной до 12—20 см) с извитым нитевидным придатком на конце. Яйца падают на каменистое дно или повисают на водорослях. Развитие продолжается 9—12 месяцев. По бокам головы развивающегося зародыша образуются нитевидные кожные выросты — наружные «жабры», которые, вероятно, облегчают всасывание желтка и, возможно, получение кислорода. Перед вылуплением эти «жабры» исчезают, и покидающая оболочку молодая химера отличается от взрослых лишь размерами. Химеры – это своеобразная группа, сочетающая признаки хрящевых и костных рыб: хрящевой скелет, но отсутствуют брызгальца. Как у костных рыб образуются жаберные крышки, но без окостенений и.т.д..
11.1
Костные рыбы являются более прогрессивной ветвью эволюции в надклассе Рыб. Это проявилось в более совершенных основных системах жизнеобеспечения (усложнились кожные покровы - космоидная, ганоидная и костная чешуя; более совершенная пищеварительная система – большая дифференциация, появляются пилорическиме выросты желудка, анальное отверстие и пр.; формируется более совершенная система дыхания – более совершенные жабры, а у части видов образуется легкие; значительно усложняется кровеносная, нервная системы – в переднем мозге двоякодышащих закладывается архипаллиум и т.д.).
Все эти перестройки отразились на особенностях образа жизни и всей биологии
Необычайно большое разнообразие форм тела рыб и различный образ их жизни определяют многообразие способов движения, которое можно свести к шести основным типам.
1. Ундулирующее (синусоидное) движение при помощи боковых ко лебательных изгибов всего тела. Такие рыбы имеют удлиненное тело, более или менее заметно сжатое с боков, особенно в его задней части, удлиненный анальный и относительно небольшой хвостовой плавники (угри, вьюны и др.). При каждом колебательном движении у головы рыбы то с той, то с другой стороны возникают вращательные (вихревые) движения воды, отталкиваясь от которых тело перемещается вперед. Коэффициент полезного действия такого движения высок, но скорости перемещения невелики. Свойственно рыбам, ведущим донный и придонный образ жизни, а также многим глубоководным видам, вынужденным при малой кормности мест их обитания экономить энергию.
2. Движение с помощью частых боковых колебательных движений задней части тела (иногда только хвостового стебля, рис. 66, внизу) свойственно большинству рыб. Они имеют более или менее компакт ное тело и мощный хвостовой стебель. Передняя часть тела выполняет функцию рассекающего воду клина, а хвостовая—движителя. По сравнению с первым типом движения уменьшается амплитуда колеба ний хвостового отдела, но резко возрастает их частота, а в результате — скорость движения. При больших скоростях работа хвостового плавника в зоне завихрений становится мало эффективной. В этой связи у быстроплавающих рыб изменяется его форма; концевые лопасти выносятся за пределы зоны завихрения.
3. Перемещение с помощью волнообразных (ундулирующих) движений одних плавников свойственно малоподвижным пелагическим и особенно придонным рыбам (скаты, камбалы). У луны-рыбы, солнечника, кузовков и некоторых других видов функцию движителей выполняют удлиненные спинной и анальный плавники, у электрических угрей — лентовидный анальный плавник (рис. 68), у морских игл — спинной плавник. У плывущих камбал ундулирующие движения окаймляющих тело спинного и анального плавников дополняются боковыми изгибаниями всего тела.
Машущие движения плавников, особенно грудных, служат добавочным движителем у многих костистых рыб при медленном плавании. Некоторые рыбы (бычки, скорпены и др.) используют грудные плавники при ползании по грунту. У морских петухов (семейство Triglidae из отряда скорпенообразных) три луча каждого грудного плав ника обособляются в пальцевидные придатки, с помощью которых рыбы ползают по дну.
Летучие рыбы из отряда сарганообразных, планируя на неподвижных, широко расставленных удлиненных парных плавниках, используемых как несущие поверхности, способны пролетать 200—400 м (с. 198). Необходимая для такого планирующего полета начальная скорость движения создается за счет энергичных движений заднего отдела тела и хвостового плавника. Мелкие длиной 3—9 см прес новодные рыбки Южной Америки из семейства клинобрюхих (Gasteropelecidae, подотряд
11.2
харциновидных из отряда карпообразных) способны выпрыгивать из воды и «пролетать» над водой 3—5 м. Выпрыгивание и полет над водой спасают от хищных рыб.
6. Особую категорию представляют так называемые паразитические формы движения. Довольно широко распространено «лоцманирование»: передвижение мелких рыб-спутников в слоях воды, увлекаемых быстро плывущей крупной рыбой или другим животным, лодкой или кораблем. Крайним примером паразитического движения могут служить рыбы прилипалы (семейство Echeneidae из отряда окунеобразных,).
Рассмотрим особенности строения и функционирования кровеносной и выделительной систем.
Кровеносная система и кровообращение. Кровеносные системы костных и хрящевых рыб сходны; различия сводятся лишь к некоторым деталям. Сердце двухкамерное (одно предсердие и один желудочек). У всех костных рыб есть венозный синус или веночная пазуха (sinus venosus), из которого кровь поступает в предсердие (atrium) и из него — в желудочек (ventriculus). У кистеперых, двоякодышащих, осетрообразных, многоперов и панцирникообразных хорошо развит артериальный конус (conus arteriosus), как и у хрящевых рыб, несущий на внутренних стенках полулунные клапаны. У амии артериальный конус сильно редуцирован, а у остальных костных рыб (Teleostei) от артериального конуса остаются лишь один-два ряда клапанов перед входом в брюшную аорту. При редукции артериального конуса образуется толстостенное вздутие начальной части брюшной аорты — луковица аорты (bulbus aortae), имеющая стенки с гладкой мускулатурой и смягчающая пульсовые толчки. Кровяное давление у большинства костных рыб несколько выше (18—120 мм рт. ст.), чем у хрящевых (7—45 мм).
Брюшная аорта (aorta ventralis), отходя от сердца, распадается на 4 пары приносящих жаберных артерий, снабжающих кровью расположенные на I—IV жаберных дугах жабры. Окислившаяся в капиллярах жаберных лепестков кровь собирается в выносящие жаберные артерии (их тоже 4 пары), которые впадают в корни спинной аорты (рис.), позади жаберного отдела сливающиеся в спинную аорту (aorta dorsalis) (рис.). Спереди корни аорты, отделив сонные артерии (рис), тоже соединяются друг с другом (рис.), образуя характерный для костных рыб головной круг. Лежащая под позвоночным столбом спинная аорта последовательно ответвляет артерии, снабжающие кровью мускулатуру и внутренние органы, и уходит в гемальный канал хвостового отдела позвоночного столба; здесь ее называют хвостовой артерией (a. Caudalis).
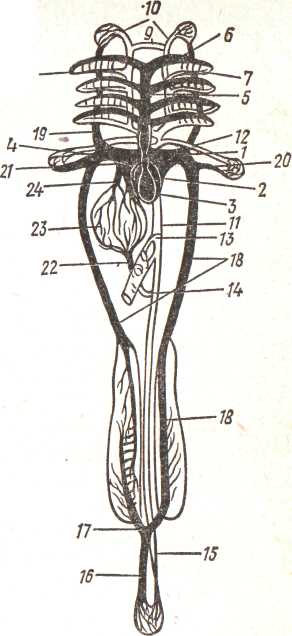
11.3
Рис. Схема кровеносной системы костистой рыбы (черным цветом показаны сосуды с венозной кровью, белым — с артериальной):
/ — венозная пазуха, 2 — предсердие, 3 — желудочек, 4 — луковица аорты, 5 — брюшная аорта, 6 — приносящие жаберные артерии, 7 — выносящие жаберные артерии, 8 — корни спинной аорты, 9 — передняя перемычка, соединяющая корни аорты, 10 — сонная артерия, // — спинная аорта, 12 — подключичная артерия, 13, —■ кишечная артерия, 14 — брыжеечная артерия, /5 — хвостовая артерия, 16 — хвостовая вена, П — воротные вены почек, IS — задняя кардинальная вена, 19 — передняя кардиналь» ная вена, 20 — подключичная вена, 21 — кювьеров проток, 22 — воротная вена печени, 23 — печень, 24 — печеночная вена
Собравшая венозную кровь из хвостового стебля хвостовая вена (vena caudalis) (рис.) входит в полость тела и разделяется на правую и левую воротные вены почек (v. portae renalis) (рис.). У части костных рыб, как и у хрящевых, они входят в почки и распадаются там на капилляры, т. е. образуют воротную систему почек; почечные капилляры, сливаясь, впадают в задние кардинальные вены. Однако у многих костистых рыб в правой почке воротная система частично редуцируется: часть крови по анастомозам сразу переходит в правую заднюю кардинальную вену, минуя капилляры. Несущие кровь из головы передние кардинальные вены (v. Cardinalis anterior) (рис. 120, 19), собирающие кровь из грудных плавников подключичные вены (v. subclavia) (рис. 120, 20) и задние кардинальные вены (v. cardinalis posterior) (рис. 120, 18) каждой стороны сливаются в соответствующий — левый или правый кювьеров проток (ductus cuvieri), впадающие в венозную пазуху. Вены, собирающие кровь от желудка, селезенки и всего кишечника, сливаются в воротную вену печени (v. portae hepatis); она входит в печень и распадается на капилляры, образуя воротную систему печени. Печеночные капилляры вновь сливаются в вены, объединяющиеся в короткую печеночную вену (v. hepatica) (рис. 120, 24), впадающую непосредственно в венозную пазуху.
В разных группах костных рыб встречаются отклонения от описанной схемы. Наиболее своеобразны особенности кровеносной системы двоякодышащих рыб, обусловленные наличием у них добавочного легочного дыхания; с ним связано возникновение легочного круга кровообращения. Предсердие свисающей со спинной стороны перегородкой (рис. 121, 5) почти полностью разделяется на две половины: в левую впадает несущая кровь из легких — легочная вена, а в правую открывается венозная пазуха. Эта перегородка в виде складки проходит через желудочек и продолжается в спиральный клапан артериального конуса. От брюшной аорты отходят 5 пар приносящих жаберных артерий (передняя из них проходит по подъязычной дуге). Через капиллярные сети жабр приносящие артерии переходят в выносящие жаберные артерии, впадающие в корни аорты; последние продолжаются вперед как сонные артерии, а позади жаберной области сливаются в спинную аорту. От последней (пятой) выносящей жаберной артерии отходит легочная артерия (рис. 121, 12), ветвящаяся в легком. При достаточном содержании кислорода в
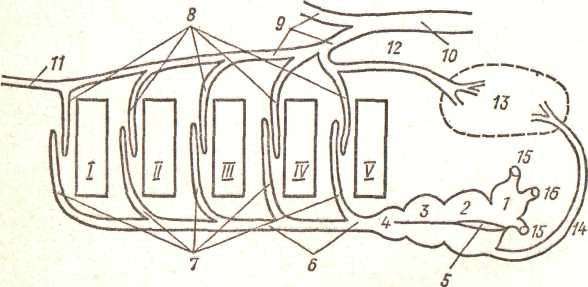
11.4
Рис. 121. Схема жаберно-легочного кровообращения двоякодышащих рыб (по Гудричу, упрощено):
I — венозная пазуха, 2 — предсердие, 3 — желудочек, 4 — артериальный конус, 5 — перегородка предсердия, переходящая в складку верхней стенки желудочка и продолжающаяся ь спиральный клапан артериального конуса, в — брюшная аорта, 7 — приносящие жаберные артерии, 8 — выносящие жаберные артерии, 9 — корни аорты, 10 — спинная аорта, // — сонная артерия, 12 — легочная артерия, 13 — легкое, 14 — легочная вена, /5 — кювьеров проток, 16 — задняя полая вена; /—V жаберные щели
воде, когда полное насыщение крови Кислородом происходит в жабрах, легкое не функционирует и получает артериальную кровь.
При неполном насыщении крови кислородом в жабрах в легкое поступает лишь частично окисленная кровь и там доокисляется. В правую половину предсердия поступает венозная кровь из венозной пазухи, в первую артериальная кровь из легкого. Перегородка предсердия затруд-1яет перемешивание крови. При сокращении предсердия более артериальная кровь течет вдоль складки желудочка и спирального клапана артериального конуса и попадает преимущественно в первые приносящие жаберные артерии, получает дополнительно кислород в жабрах и идет в голову и корни аорты. Более венозная кровь из правой части предсердия идет преимущественно в задние приносящие жаберные артерии, получает в жабрах какое-то количество кислорода и попадает в корни аорты, а частично — через легочную артерию — легкие. Большой и малый круги кровообращения, таким образом, разобщены очень слабо. Однако это делает возможным одновременное использование жаберного и легочного дыхания; причем более артериальная кровь поступает в голову, а по телу и в легкие идет смешанная кровь. Есть своеобразные черты и в периферической системе. Задние ординальные вены развиваются относительно слабо. Основная масса крови из капилляров обеих почек в конечном счете попадает в крупный сосуд — заднюю полую вену (v. cava posterior), впадающую непосредственно в венозную пазуху (рис. 121, 16).
Таким образом, у двоякодышащих появляются изменения, которые напоминают особенности кровеносной системы земноводных.
Можно предполагать, что у имевших легкие кистеперых — предков земноводных ■— кровеносная система была сходна с кровеносной системой ныне живущих двоякодышащих. Поэтому изучение становления и функционирования кровеносной системы двоякодышащих позволяет представить возможные пути морфологических перестроек, сопровождавших обособление первого класса наземных позвоночных — земноводных — от рыбообразных предков.
Основные показатели крови (кислородная емкость, количество гемоглобина и эритроцитов, ионный состав) обычно заметно выше у более подвижных пелагических видов и меньше — у малоподвижных донных форм. В пределах класса костных рыб общее количество крови варьирует от 1,1 до 7,3% от массы тела, число эритроцитов — от 580 тыс. до 4,1 млн, в 1 мм3, содержание гемоглобина изменяется пределах 1,1—17,4 г% и 0,5—3,4 г на 1 кг массы; кислородная емкость крови составляет 1,5—23,0 объемных процента. Верхняя граница этих показателей у костных рыб заметно выше, чем у хрящевых рыб (см. табл. 5, с. 156). Живущие в водах Антарктиды белокровные рыбы (около 10 видов сем. Chaenichthyidae, подотр. нототениевые, отр. окунеобразные) в отличие от других рыб не имеют эритроцитов и гемоглобина. Кровь у них бесцветная. Органами дыхания служат не столько жабры, сколько обильно снабженная капиллярами кожа (до 45 мм длины капилляров на 1 мм2 поверхности тела). Поверхность капилляров кожи и плавников в 2 раза превышает общую поверхность тела. Живут эти рыбы в воде с высоким содержанием кислорода и постоянно низкой
11.5
температурой (ниже +2° С). Отличаются большими размерами сердца и низким содержанием кислорода в крови.
Важную роль в обеспечении энергетики организма играют углеводы, циркулирующие в крови (глюкоза, гликоген и др.). В отличие от высших позвоночных у рыб колебания сахара в крови очень велики, что связано с несовершенным механизмом регуляции. Выявляются сезонные изменения: повышение Сахаров в периоды размножения и миграций (обычно в теплое время года) и снижение на зимовках, при малоподвижном образе жизни. Регуляция обеспечивается использованием запасов гликогена в печени под контролем гормональной и нервной систем, т. е. обычным путем сезонных перестроек метаболизма. У высокоподвижных и активных рыб (тунцы и др.) содержание сахара в крови удерживается в пределах 60—90 мг% (у осетровых поднимается до 115 мг%), а у малоподвижных донных рыб (морской чёрт) обычно не бывает выше 10 мг% (у хрящевых — 20—50 мг%). Кровь костных рыб отличается от крови хрящевых рыб большим содержанием белков в плазме. Образование форменных элементов крови происходит преимущественно в селезенке и в почках (особенно в их передних отделах).
Органы выделения и водно-солевой обмен. Система органов выделения, основу которой у всех рыб составляют почки, не только выводит продукты распада азотистых веществ, но и обеспечивает физико-химическую устойчивость внутренней среды организма: осмотического давления, кислотно-щелочного ионного равновесия. В этих процессах участвуют почки, жаберный аппарат, кожа, пищеварительная трубка и печень. Эти системы различаются у хрящевых и костных рыб.
Парные мезонефрические (туловищные) почки костных рыб лежат ниже позвоночного столба, почти вдоль всей полости тела. У многих видов в задней половине правая и левая почки сливаются вместе. Вольфовы каналы выполняют функцию мочеточников. По выходе из почки мочеточники правой и левой почек сливаются и, образовав тонкостенный полый вырост — мочевой пузырь (vesica urinaria) , открываются наружу мочевым отверстием (у двоякодышащих — в клоаку). Почки получают артериальную кровь из почечных артерий; большие порции венозной крови приходят сюда по воротным венам почек из хвостового отдела. У пресноводных рыб хорошо развиты боуменовы капсулы с крупными клубочками. У морских костных рыб размеры клубочков уменьшаются, а у нескольких десятков видов из разных отрядов (жабы-рыбы, некоторые морские иглы и др.) клубочки практически редуцированы (агломерулярные почки).
У подавляющего большинства лучеперых рыб, особенно у костистых, конечным продуктом распада азотистых (в том числе и белковых) соединений, выводимым из организма, служит аммиак (как и у большинства беспозвоночных). Это парадоксальное явление находит свое объяснение в том, что жаберные лепестки этих рыб способны интенсивно выделять в окружающую среду аммиак и ионы солей, обеспечивая таким образом значительную часть процессов выведения продуктов распада из организма. Это объясняет, почему у многих видов почки особенно их верхние части) потеряли исходную функцию, превратившись в лимфоидный орган, участвующий в кроветворении. Часть азота выводится в виде мочевины, триметиламиноксида, креатина и близких соединений; она возрастает у морских видов по сравнению с пресноводными. Двоякодышащие рыбы, впадающие в оцепенение при высыхании водоемов (протоптерус), в активном состоянии выделяют аммиак, а в оцепенении — мочевину, накапливающуюся в организме. Она выводится после пробуждения рыбы (напомним: аммиак много токсичнее мочевины.)
Выделение продуктов распада у рыб тесно связано с водно-солевым обменом, обеспечивающим осмотическое и кислотно-щелочное равновесие в тканях. У морских и пресноводных рыб эти процессы протекают различно. Пресноводные рыбы
11.6
живут в гипотонической реде (осмотическое давление в их тканях значительно выше осмоти-еского давления пресной воды (табл. 6, с. 158). Поэтому вода постоянно проникает в организм через кожу (рис. 123, /), жабры и с пищей. По некоторым данным за 3—5 ч поступающей извне водой заменяется вся вода организма; ему постоянно грозит излишнее «обводнение», высоко развитый фильтрационный аппарат почек позволяет выводить избытки воды; за сутки пресноводные рыбы выделяют от 50 до 300 мл очи и более на 1 кг массы тела. Потери солей при выведении больших количеств мочи компенсируются активной реабсорбцией солей в почечных канальцах и поглощением солей жабрами из внешней среды.
12.1