

Поведение и внд
№ 4 Торможение в коре головного мозга, его значение и виды. Характеристика внешнего и внутреннего торможения и современные представления об их механизмах.
Торможение в коре головного мозга, его значение и виды:
В зависимости от условий возникновения коркового торможения различают две формы – безусловное (внешнее) и условное (внутреннее).
При исследовании взаимоотношений возбуждения и торможения в коре мозга было установлено, что в течение нескольких секунд после воздействия тормозного раздражителя эффект положительных условных раздражителей усиливается. И наоборот, после применения положительных условных раздражителей усиливается действие тормозящих раздражении.
Внешнее (безусловное) торможение наблюдается в случаях, когда на животное с ранее выработанным условным рефлексом неожиданно действует какой-то новый, довольно сильный внешний раздражитель. Внешнее торможение проявляется также в новой обстановке. В этом случае у животного возникает ориентировочно-исследовательская деятельность, которая и является причиной торможения ранее выработанного условного рефлекса. Внешнее торможение не требует обучения. Торможение происходит в результате взаимодействия двух возбуждений. Возбуждение, обусловленное ориентировочно-исследовательской реакцией, оказывается более сильным и затормаживает более слабое возбуждение. С системных позиций при этом более сильная функциональная система на уровне отдельных нейронов мозга вытормаживает более слабую.
Внутреннее (условное) торможение возникает в самой дуге условного рефлекса. Требует специального обучения. Главным условием формирования внутреннего торможения является отсутствие подкрепления условного сигнала. При этом у животных сначала, так же как и в случае внешнего торможения, в ответ на условный раздражитель развивается ориентировочно-исследовательская реакция, которая в дальнейшем сменяется отрицательной эмоцией.
Различают несколько видов внутреннего торможения:
Угасательное торможение развивается в тех случаях, когда условный сигнал ранее выработанного условного рефлекса перестает подкрепляться. Например, пищевой условный раздражитель при его последовательных предъявлениях перестает подкрепляться пищей. В этом случае величина слюноотделения на каждый последующий условный сигнал постепенно уменьшается и в конце концов достигает нулевых значений.
Дифференцировочное торможение формируется в случае, когда один из условных раздражителей подкрепляется, а другой, близкий к нему по физическим параметрам, не подкрепляется. Торможение проявляется в этом случае по отношению к неподкрепляемому воздействию и развивается в две фазы. Сначала возникает фаза генерализации, в которой животное отвечает на оба условных — подкрепляемый и неподкрепляемый — раздражителя. Затем формируется стадия концентрации, когда на ранее подкрепляемый условный раздражитель животное отвечает условнорефлекторной реакцией, а на неподкрепляемый условный раздражитель условнорефлекторная реакция не проявляется.
Запаздывательное торможение формируется в случаях, когда подкрепление отставлено от условного раздражителя, например, на 2—3 мин. При этом при пищевых запаздывательных условных рефлексах торможение проявляется в течение всего времени действия условного сигнала.
Условный тормоз проявляется в тех случаях, когда условный раздражитель подкрепляется, а сочетание его с другим условным раздражителем не подкрепляется. В этом случае второй условный раздражитель становится тормозом.
Механизм внутреннего торможения. Павлов считал, что внутреннее торможение развивается в корковых клетках представительства условных раздражителей. Однако опыты с регистрацией вегетативных компонентов условнорефлекторной деятельности Анохина свидетельствуют о том, что при торможении слюноотделения при пищевом условном рефлексе изменения сердцебиений и дыхания продолжают регистрироваться.
вкусовых раздражителей на боковые поверхности языка.
№ 10 Архитектоника функциональной системы целенаправленного поведенческого акта. Исполнительные механизмы системной организации поведенческого акта с позиции теории функциональной системы П.K.Анохина и их характеристики.
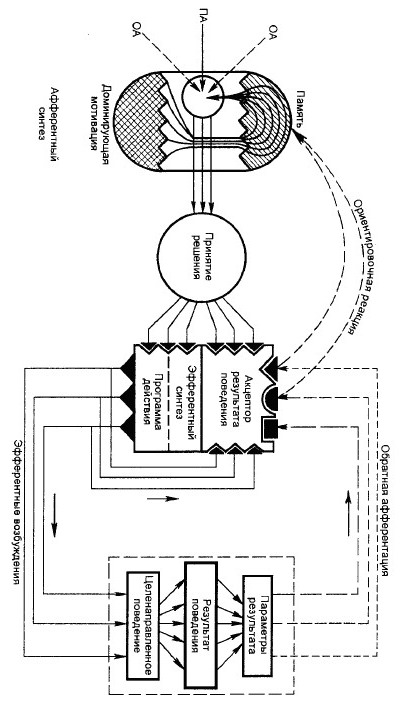
ОА – обстановочная афферентация; ПА – пусковая афферентация.
Центральная архитектоника поведенческого акта строится деятельностью головного мозга, являясь атрибутом сложных динамических корково-подкорковых взаимоотношений.
Первой, инициативной стадией центральной архитектоники поведенческого акта является стадия афферентного синтеза, которая состоит из нескольких компонентов.
Ведущим компонентом является доминирующая биологическая мотивация, которая строится на основе нервно-гуморальной сигнализации различными метаболическими потребностями.
Доминирующие биологические мотивации голода, страха, жажды, полового возбуждения и др. за счет восходящих активирующих влияний специальных гипоталамических центров избирательно охватывают различные отделы головного мозга, включая кору. Биологические мотивации могут самостоятельно сформировать поведенческий акт. При этом внешние факторы играют роль ключевых, раскрывающих в определенных условиях генетические механизмы поведенческих актов.
Влияния внешней среды составляют второй компонент афферентного синтеза — обстановочную афферентацию, которая непрерывно поступает в ЦНС при действии разнообразных факторов внешней среды на многочисленные экстерорецепторы живых организмов.
Соотношения доминирующей мотивации и обстановки динамичны, они строятся по принципу доминанты — в первую очередь удовлетворяются биологические или обстановочные воздействия, наиболее значимые для выживания или социальной адаптации.
Третьим компонентом афферентного синтеза является память. Прежде всего это генетическая память, к которой в построении поведения постоянно адресуются врожденные биологические мотивации. Механизмы памяти при определенных условиях могут самостоятельно сформировать поведенческий акт или существенно повлиять на его организацию.
№ 3 Условные и безусловные рефлексы. Черты их сходства и различия, значение для приспособительной деятельности организма. Методики, условия и правила выработки условных рефлексов. Механизмы о6разования временных связей (И.П. Павлов, П.К. Анохин).
Условные и безусловные рефлексы: смотри №102
Правила выработки условных рефлексов:
1. Наличие у животного потребности и соответствующей мотивации. Например, в случае выработки пищевого условного рефлекса животное должно быть голодным. При формировании оборонительного условного рефлекса животное в ответ на повреждающее безусловное воздействие должно испытывать страх.
2. Условный раздражитель должен обязательно подкрепляться безусловным, т.е. удовлетворением жизненно важной потребности.
3. Условный раздражитель должен предшествовать подкреплению.
4. Условный раздражитель должен восприниматься животным, т.е. первоначально вызывать ориентировочно-исследовательскую деятельность. Условный раздражитель должен нести в себе экологическую значимость для животного, восприниматься им. Так, у рыб в качестве условных раздражителей более адекватными являются движение воды, изменение ее состава и др. Для высших животных это могут быть световые, звуковые, обонятельные и другие сенсорные раздражители.
5. Подкрепление по своей биологической значимости и силе должно быть сильнее условного раздражителя.
6. Условный раздражитель должен неоднократно сочетаться с безусловным подкреплением.
7. При выработке соответствующего условного рефлекса у животных должны отсутствовать конкурирующие мотивации. Например, в случае выработки пищевого условного рефлекса у животного не должен быть переполнен мочевой пузырь или животное не должно подвергаться каким-либо повреждающим воздействиям.
8. Наконец, субъект, у которого вырабатываются условные рефлексы, должен быть здоров.
Механизмы образования временных:
Павлов связывал образование условных рефлексов главным образом с деятельностью коры большого мозга, хотя он не отрицал участия в этом процессе и ближайших подкорковых образований. Он рассматривал процесс образования условного рефлекса как взаимодействие двух дуг возбуждений: дуги условного и безусловного рефлексов. Между этими дугами, при повторных сочетаниях образуется временная связь.
Павлов обозначил ее «временной», так как при отсутствии подкрепления она быстро разрушается и условный рефлекс исчезает. Временная связь между условным раздражением и подкреплением формируется в коре большого мозга между пунктами (очагами) представительства условного сигнала и безусловного подкрепления. Образованию условнорефлекторной временной связи в коре большого мозга способствуют доминантные отношения. При этом корковый «очаг» безусловного подкрепления, будучи доминантным, притягивает к себе возбуждения, ранее вызванные условным раздражителем. Именно эти свойства способствуют образованию временной связи между пунктами условного и безусловного раздражений коры больших полушарий. Вследствие этого условный раздражитель начинает вызывать условнорефлекторный ответ.
№ 3 Условные и безусловные рефлексы. Системная организация врожденного и приобретенного поведения.
Павлов предложил рассматривать два вида поведенческих рефлексов — безусловные и условные. Безусловные рефлексы— врожденные. Безусловные рефлексы возникают на основе врожденных рефлекторных дуг. При действии адекватных раздражителей на соответствующие рецепторы безусловные рефлексы проявляются относительно постоянно.
К сложным безусловным рефлексам относятся пищевые, оборонительные, половые, ориентировочно-исследовательские, родительские и др. Следует особо выделить ориентировочно-исследовательскую деятельность — реакцию животных на неожиданные, как правило, новые раздражители. Сложные безусловные рефлексы проявляются в виде специфических поведенческих реакций животных при действии на них соответствующих раздражителей. Наиболее демонстративен в этом плане сложный пищевой рефлекс. Он проявляется при действии пищи на дистантные рецепторы или на рецепторы пищеварительного тракта животного в двигательной, а также секреторной и других вегетативных реакциях — изменении дыхания, деятельности сердца и др. Сложный оборонительный рефлекс наряду с двигательной реакцией животного включает также изменение ряда вегетативных функций: секреторной деятельности пищеварительных желез, деятельности сердца, дыхания, потоотделения и т.д.
Условный рефлекс — приобретаются живыми существами в индивидуальной жизни. Они связаны с обучением. Это чрезвычайно изменчивая форма рефлекторной деятельности. Ответное действие животного определяется не самим стимулом, а возникает в результате неоднократного совпадения того или иного внешнего (условного) стимула с жизненно важной деятельностью (безусловными рефлексами). Тогда ранее относительно индифферентный стимул начинает опережающе вызывать реакцию, характерную для безусловного раздражителя. Иными словами, в выработанном условном рефлексе условный стимул опережающе отражает свойства сочетанного с ним безусловного раздражителя.
Непременным условием образования условных рефлексов является подкрепление, когда ранее индифферентный раздражитель неоднократно сочетается с последующим безусловным рефлексом.
Другой принцип, характеризующий условнорефлекторную деятельность – принцип сигнальности. Ответная реакция организма при действии не него раздражитеоя несет в себе свойства будущего безусловного воздействия. Условный раздражитель сигнализирует о последующие безусловном рефлексе.
Условные рефлексы классифицируют:
- по названию условных раздражителей — световые, звуковые, обонятельные, тактильные;
- по названию анализатора, воспринимающего условный раздражитель,— зрительные, слуховые, кожные;
- по характеру подкрепления — пищевые, оборонительные, половые;
- по методу выработки — коротко- и длительноотставленные, запаздывательные, следовые и совпадающие.
При короткоотставленных условных рефлексах интервал между условным раздражителем и подкреплением обычно равен 10—20 с и не превышает 30 с. В длительноотставленных условных рефлексах этот интервал составляет более 30 с. В запаздывательных условных рефлексах интервал между условным сигналом и подкреплением равен 3 мин. В следовых условных рефлексах подкрепление предоставляется животному после прекращения действия условного раздражителя. При совпадающих условных рефлексах условный сигнал и подкрепление предоставляются животному одновременно.
№ 4 Торможение в коре головного мозга, его значение и виды. Характеристика внешнего и внутреннего торможения и современные представления об их механизмах.
Торможение в коре головного мозга, его значение и виды:
В зависимости от условий возникновения коркового торможения различают две формы – безусловное (внешнее) и условное (внутреннее).
При исследовании взаимоотношений возбуждения и торможения в коре мозга было установлено, что в течение нескольких секунд после воздействия тормозного раздражителя эффект положительных условных раздражителей усиливается. И наоборот, после применения положительных условных раздражителей усиливается действие тормозящих раздражении.
Внешнее (безусловное) торможение наблюдается в случаях, когда на животное с ранее выработанным условным рефлексом неожиданно действует какой-то новый, довольно сильный внешний раздражитель. Внешнее торможение проявляется также в новой обстановке. В этом случае у животного возникает ориентировочно-исследовательская деятельность, которая и является причиной торможения ранее выработанного условного рефлекса. Внешнее торможение не требует обучения. Торможение происходит в результате взаимодействия двух возбуждений. Возбуждение, обусловленное ориентировочно-исследовательской реакцией, оказывается более сильным и затормаживает более слабое возбуждение. С системных позиций при этом более сильная функциональная система на уровне отдельных нейронов мозга вытормаживает более слабую.
Внутреннее (условное) торможение возникает в самой дуге условного рефлекса. Требует специального обучения. Главным условием формирования внутреннего торможения является отсутствие подкрепления условного сигнала. При этом у животных сначала, так же как и в случае внешнего торможения, в ответ на условный раздражитель развивается ориентировочно-исследовательская реакция, которая в дальнейшем сменяется отрицательной эмоцией.
Различают несколько видов внутреннего торможения:
Угасательное торможение развивается в тех случаях, когда условный сигнал ранее выработанного условного рефлекса перестает подкрепляться. Например, пищевой условный раздражитель при его последовательных предъявлениях перестает подкрепляться пищей. В этом случае величина слюноотделения на каждый последующий условный сигнал постепенно уменьшается и в конце концов достигает нулевых значений.
Дифференцировочное торможение формируется в случае, когда один из условных раздражителей подкрепляется, а другой, близкий к нему по физическим параметрам, не подкрепляется. Торможение проявляется в этом случае по отношению к неподкрепляемому воздействию и развивается в две фазы. Сначала возникает фаза генерализации, в которой животное отвечает на оба условных — подкрепляемый и неподкрепляемый — раздражителя. Затем формируется стадия концентрации, когда на ранее подкрепляемый условный раздражитель животное отвечает условнорефлекторной реакцией, а на неподкрепляемый условный раздражитель условнорефлекторная реакция не проявляется.
Запаздывательное торможение формируется в случаях, когда подкрепление отставлено от условного раздражителя, например, на 2—3 мин. При этом при пищевых запаздывательных условных рефлексах торможение проявляется в течение всего времени действия условного сигнала.
Условный тормоз проявляется в тех случаях, когда условный раздражитель подкрепляется, а сочетание его с другим условным раздражителем не подкрепляется. В этом случае второй условный раздражитель становится тормозом.
Механизм внутреннего торможения. Павлов считал, что внутреннее торможение развивается в корковых клетках представительства условных раздражителей. Однако опыты с регистрацией вегетативных компонентов условнорефлекторной деятельности Анохина свидетельствуют о том, что при торможении слюноотделения при пищевом условном рефлексе изменения сердцебиений и дыхания продолжают регистрироваться.
№ 4 Торможение условнорефлекторной деятельности. Безусловное торможение и его виды. Условное торможение и его виды. Механизмы безусловного и условного торможения.
Условнорефлекторная и временная связь в головном мозге, так же как и приобретаемые в индивидуальной жизни программы поведения, довольно динамичны. Они становятся прочными при длительных однотипных способах удовлетворения соответствующих потребностей организма в стабильных условиях внешней среды. В то же время при специальных воздействиях они нарушаются, исчезают ранее выработанные условные рефлексы.
Процессы разрушения временных связей происходят в результате торможения условнорефлекторной деятельности. Выявлено несколько разновидностей торможения условных рефлексов.
Внешнее (безусловное) торможение наблюдается в случаях, когда на животное с ранее выработанным условным рефлексом неожиданно действует какой-то новый, довольно сильный внешний раздражитель. Внешнее торможение проявляется также в новой обстановке. В этом случае у животного возникает ориентировочно-исследовательская деятельность, которая и является причиной торможения ранее выработанного условного рефлекса. Внешнее торможение не требует обучения. Торможение происходит в результате взаимодействия двух возбуждений. Возбуждение, обусловленное ориентировочно-исследовательской реакцией, оказывается более сильным и затормаживает более слабое возбуждение. С системных позиций при этом более сильная функциональная система на уровне отдельных нейронов мозга вытормаживает более слабую.
Внутреннее (условное) торможение возникает в самой дуге условного рефлекса. Требует специального обучения. Главным условием формирования внутреннего торможения является отсутствие подкрепления условного сигнала. При этом у животных сначала, так же как и в случае внешнего торможения, в ответ на условный раздражитель развивается ориентировочно-исследовательская реакция, которая в дальнейшем сменяется отрицательной эмоцией.
Различают несколько видов внутреннего торможения:
Угасательное торможение развивается в тех случаях, когда условный сигнал ранее выработанного условного рефлекса перестает подкрепляться. Например, пищевой условный раздражитель при его последовательных предъявлениях перестает подкрепляться пищей. В этом случае величина слюноотделения на каждый последующий условный сигнал постепенно уменьшается и в конце концов достигает нулевых значений.
Дифференцировочное торможение формируется в случае, когда один из условных раздражителей подкрепляется, а другой, близкий к нему по физическим параметрам, не подкрепляется. Торможение проявляется в этом случае по отношению к неподкрепляемому воздействию и развивается в две фазы. Сначала возникает фаза генерализации, в которой животное отвечает на оба условных — подкрепляемый и неподкрепляемый — раздражителя. Затем формируется стадия концентрации, когда на ранее подкрепляемый условный раздражитель животное отвечает условнорефлекторной реакцией, а на неподкрепляемый условный раздражитель условнорефлекторная реакция не проявляется.
Запаздывательное торможение формируется в случаях, когда подкрепление отставлено от условного раздражителя, например, на 2—3 мин. При этом при пищевых запаздывательных условных рефлексах торможение проявляется в течение всего времени действия условного сигнала.
Условный тормоз проявляется в тех случаях, когда условный раздражитель подкрепляется, а сочетание его с другим условным раздражителем не подкрепляется. В этом случае второй условный раздражитель становится тормозом.
Механизм внутреннего торможения. Павлов считал, что внутреннее торможение развивается в корковых клетках представительства условных раздражителей. Однако опыты с регистрацией вегетативных компонентов условнорефлекторной деятельности Анохина свидетельствуют о том, что при торможении слюноотделения при пищевом условном рефлексе изменения сердцебиений и дыхания продолжают регистрироваться.
№ 5 Типы высшей нервной деятельности. Типологические особенности ВНД человека (Гиппократ, И.П.Павлов), характеристика художественного, мыслительного и среднего типов психики человека.
Учение о типах поведения (темпераментах) восходит еще к Гиппократу, который по преобладанию той или иной внутренней среды подразделял людей на сангвиников, холериков, меланхоликов и флегматиков.
Павлов в основу классификации типов ВНД у животных положил следующие свойства основных нервных процессов возбуждения и торможения: силу, уравновешенность и подвижность.
По силе нервных процессов животных подразделяют на сильных и слабых. При этом сила процесса возбуждения характеризует предел работоспособности животных, за которым наступает пессимальное торможение.
Уравновешенность отражает баланс между процессами возбуждения и торможения.
Подвижность отражает способность смены процесса возбуждения на торможение и наоборот.
Типы высшей нервной деятельности у собак. Павлов соотнес указанные свойства нервной системы с классификацией темпераментов по Гиппократу.
Сангвиник — сильный по процессам возбуждения и торможения, уравновешенный, подвижный.
Холерик — сильный, неуравновешенный (возбудимый), подвижный.
Флегматик — сильный, уравновешенный, инертный.
Меланхолик — слабый.
Системная классификация типов высшей нервной деятельности. Системный подход изменил представления о типах ВНД человека и животных. Классификация типов при этом строится с учетом индивидуальных особенностей формирования у субъектов стадий афферентного синтеза, принятия решения, способности предвидения и оценки результатов действия.
Значение различных стадий системной организации поведенческих актов меняется у одних и тех же индивидов. Человек может хорошо принимать решения в одной ситуации и быть совершенно беспомощным в другой.
Индивидуальные особенности организации различных функциональных систем у каждого индивида должны учитываться при разработке и применении фармакологических и нелекарственных средств.
Павлов ввел представление о двух сигнальных системах действительности. Первая сигнальная система связана с физическими свойствами условных раздражений. Она присуща животным и человеку. Вторая сигнальная система связана со словесными воздействиями на человека. Павлов рассматривал слово как «сигнал сигналов».
Классификация типов высшей нервной деятельности у человека. По преобладанию первой и второй сигнальной систем Павлов подразделял людей на мыслительные, художественные и смешанные типы. У мыслительных типов преобладает вторая сигнальная система, у художественных типов — первая.
№ ? Системные механизмы боли. Классификация болей. Нейрофизиологические и биохимические компоненты боли. Биологическое значение боли.
Боль является интегративной функцией организма, которая мобилизует организм и его разнообразные функциональные системы на защиту от воздействующих вредящих факторов и включает такие компоненты, как сознание, ощущение, память, мотивации, вегетативные, соматические, поведенческие реакции, эмоции. Боль может возникать при сильном раздражении различных сенсорных рецепторов: температуры, давления и т.д.
Боль разделяют на два типа. Первый тип — острая, «эпикритическая», боль, которая быстро осознается, легко детерминируется и локализуется, к ней быстро развивается адаптация, и она продолжается не дольше, чем действие стимула. Второй тип — тупая, «протопатическая», боль, которая осознается более медленно, плохо локализуется, сохраняется длительное время и не сопровождается развитием адаптации. Считается, что второй тип боли эволюционно более древний и менее совершенный как сигнал опасности.
Ощущение боли можно классифицировать по качествам, определяемым либо по месту ее возникновения, либо по характеру. В частности, разделяют боль соматическую и висцеральную. В свою очередь соматическая боль состоит из двух подклассов: поверхностной и глубокой боли. Соматическая боль, возникающая в коже, называется поверхностной, тогда как боль, исходящая от мышц, костей, суставов или соединительной ткани, получила название глубокой боли. Самым известным примером глубокой боли является головная боль. По времени формирования болевого ощущения боль разделяют на раннюю и позднюю.
Висцеральная боль сходна с глубокой болью тем, что сопровождается такими же вегетативными реакциями.
Особые формы боли. Кардиогенная боль, источником которой является главным образом ишемия миокарда, возникает вследствие недостаточности коронарного кровообращения. Образующиеся при этом биологически активные вещества (брадикинин) и продукты метаболизма раздражают нервные окончания.
Среди нейрогенных болей выделяют лицевые боли, обусловленные невралгией черепного нерва или симпаталгией. Фантомные боли появляются после ампутации конечности. Гемиалгии — жестокие, трудно переносимые боли в половине тела, связаны с раздражением каким-либо патологическим процессом (опухоли, сосудистые заболевания, инсульты) зрительного бугра.
Каузалгии («жгучие боли») — возникают при частичном повреждении нерва с неполным нарушением проводимости и явлениями раздражения вегетативных волокон. К особым формам относят проецируемую боль, т.е. состояние, при котором место, на которое действует повреждающий стимул, не совпадает с тем, где эта боль ощущается.
Нейрофизиологические компоненты боли:
Участие в механизмах болевого возбуждения хемоноцицепторов предполагает вовлечение в эти процессы химических веществ. Основными химическими веществами, которые вызывают активацию хемоноцицепторов, являются прежде всего медиаторы. Установлено, что ацетилхолин, норадреналин, серотонин, а также некоторые химические элементы, как, например, калий, изменяющий возбудимость мембраны нервной клетки, вызывают болевые ощущения. Кроме того, имеются вещества, которые, не являясь медиаторами, усиливают возбуждение хемоноцицепторов, на основе которого возникает болевое ощущение. Эти вещества постоянно присутствуют в тканях, но при нарушении окислительных процессов их образование резко увеличивается. При нарушении целостности или функционального состояния тканей (травма, воспаление) увеличивается образование хлорида калия, гистамина, серотонина, простагландинов, кининов, например брадикинина и вещества П, повышающих возбудимость как механо-, так и хемоноцицепторов.
Модуляторное усиливающее действие вещества П на передачу ноци-цептивной импульсации отмечено на уровне нейронов задних рогов спинного мозга.
Существует ряд химических веществ, которые в большей или меньшей степени участвуют в механизмах формирования ноцицептивного возбуждения на разных уровнях ЦНС. К ним относятся модуляторы и медиаторы ноцицептивной импульсации, модуляторы и медиаторы возбуждения или торможения образований мозга, формирующих боль.
Значение боли: существование в организме ноцицептивной системы, формирующей болевые ощущения, направлено на устранение воздействий, которые могут причинить организму вред, угрожая жизнедеятельности.
№ 7 Физиология эмоций. Эмоции как компонент системной архитектоники поведенческих актов. Теории эмоций. Эмоциональный стресс. Профилактика эмоционального стресса.
Эмоции – субъективное переживание человеком своего внутреннего состояния, в частности потребностей, а также социальных факторов окружающей среды.
Системные механизмы эмоций: теория функциональных систем рассматривает эмоциональные состояния в отрезке поведенческой деятельности, включающем доминирующую потребность и ее удовлетворение, в системном кванте поведения. В этом отрезке поведенческой деятельности эмоции определяют субъективную оценку потребности, предвидение свойств потребного результата и, наконец, оценку действия на организм подкрепляющих раздражений.
С позиций теории функциональных систем эмоциональная окраска поведенческого акта определяется отношением субъекта к доминирующей потребности и достигнутому результату. В случае достижения потребного приспособительного результата возникает эмоция положительного качества, биологический и социальный смысл которой — санкционировать успех поиска. В случае отсутствия достижения приспособительного результата на основе ориентировочно-исследовательской реакции нарастает общая отрицательная эмоция неудовлетворенности, которая позволяет субъекту преодолевать препятствия или строить другие формы результативной деятельности. При длительном отсутствии возможности достижения потребного результата возникает эмоциональный стресс. На основе предшествующих удовлетворений ведущих биологических и социальных потребностей эмоции все в большей степени включаются в формирование аппарата предвидения потребного результата — акцептора результата действия, определяя формирование эмоционально окрашенной цели. Эмоции, следовательно, зависят от обучения.
Теории эмоций:
Теория подкорковых центров: связывает генез эмоций с глубинными структурами мозга.
Многочисленные клинические наблюдения указывают на то, что при локализации очага повреждения, например опухоли или кровоизлияния, в глубинных структурах мозга у пациентов наблюдаются нарушения эмоциональных переживаний от повышенной раздражительности до полного подавления эмоций.
Корковая теория эмоций. Эксперименты с удалением у животных коры головного мозга отчетливо продемонстрировали, что эмоции у животных после удаления коры мозга сохраняются, но изменяют свой характер. Как правило, у таких животных эмоциональные реакции на внешние раздражители, особенно агрессивные, становятся более выраженными, яркими.
Периферическая теория эмоций: в формировании эмоций существенная роль принадлежит влияниям, идущим в ЦНС со стороны внутренних органов.
Интегративная (корково-подкорковая) теория эмоций: эмоции являются целостной реакцией мозга, формирующейся на основе объединения (интеграции) различных структур мозга: подкорковых образований и коры.
Эндогенные эмоции. Эмоциональные возбуждения возникают первично в мотивациогенных центрах гипоталамуса, а затем генерализованно распространяются в восходящем направлении на лимбические структуры и кору большого мозга. Таким путем формируются ощущения потребностей голода, жажды, страха, полового возбуждения.
Экзогенные эмоции. Эмоциональные реакции возникают под первичным влиянием внешних воздействий. В этом случае возбуждения, вызванные действием на организм внешних факторов, первично по специфическим сенсорным путям достигают клеток соответствующих проекционных зон коры большого мозга и активируют корковые механизмы памяти. Только после этого возбуждения распространяются в нисходящем направлении на эмоциогенные подкорковые и, в частности, лимбические центры, формируя в зависимости от внешних воздействий и следов памяти в одних случаях положительные, а в других — отрицательные эмоциональные реакции субъекта.
Профилактика эмоционального стресса:
1. интенсивная (дозированная) мышечная нагрузка
2. микроволновое воздействие – поле УВЧ, СВЧ, определенной частоты модуляции и импульсные токи.
3. переключение на любимую работу, от которой человек получает положительные эмоции. Хобби.
4. Плавание, сауна, прием антиоксидантов, некоторых олигопептидов (вещество П, пептид, вызывающий дельта сон, бета – эндорфин, пролактин)
№ 11 Физиология памяти. Виды памяти. Современные представления о механизмах памяти. Характеристика краткосрочной и долгосрочной памяти. Память как компонент системной архитектоники поведенческих актов.
Память — способность живых существ запоминать, сохранять и воспроизводить информацию о ранее воздействовавших на них событиях. Память тесно связана с обучением. С точки зрения системной организации физиологических функций, память присуща всем компонентам системной архитектоники поведенческого акта — стадии афферентного синтеза, принятия решения, акцептора результата действия и эфферентного синтеза.
Виды памяти. Выделяют три вида памяти: кратковременную, промежуточную и долговременную.
Кратковременная память определяет значимость поступающей информации для организма. Если эта информация важна для организма, особенно для удовлетворения его ведущих потребностей, она затем обрабатывается в промежуточной памяти и переходит в долговременную память. В противном случае она быстро забывается.
Промежуточная память определяет сохранение полученной организмом информации в течение нескольких минут или часов. Такая память определяет, например, формирование мыслей при разговоре, запоминание адресов, телефонных разговоров, некоторых заданий (так называемая рабочая память).
Долговременная память сохраняется всю жизнь. Важные для субъекта, особенно эмоционально окрашенные события, запечатляются в долговременной памяти достаточно ярко.
Особую форму представляет эмоциональная память — запечатление ярких положительных и отрицательных эмоциональных переживаний.
Процесс памяти включает 4 стадии:
- восприятие, запечатление, запоминание информации;
- хранение информации;
- воспроизведение необходимой информации;
- забывание.
Кратковременная память формируется на основе непосредственно сенсорного отпечатка внешнего мира. При этом в памяти уже удерживается ограниченная, выделенная доминирующей мотивацией информация о внешней среде, способствующая удовлетворению ведущей потребности организма. Кратковременная память позволяет в течение нескольких секунд или минут удерживать и воспроизводить отобранную часть информации.
Процессы последующей за кратковременной промежуточной памяти обычно разыгрываются в течение нескольких часов после обучения. В этот период времени экстремальные механические и химические воздействия способны стереть память, но по истечении 4 ч следы кратковременной памяти становятся устойчивыми. Происходит консолидация памяти.
Кратковременная память нарушается при таких воздействиях на организм, как электрошок, сильные мозговые травмы, судороги, наркоз, гипоксия. При этом наблюдается ретроградная амнезия — потеря памяти на события, предшествовавшие воздействию. Установлено, что объем кратковременной памяти человека составляет 7±2 единицы, т.е. бессмысленные слова после однократного их применения воспроизводятся испытуемыми лишь в эквивалентном количестве.
В основе современных представлений о механизмах кратковременной памяти лежит несколько гипотез: корково – подкорковая реверберация возбуждений, синаптическая теория.
Долговременная память определяет сохранение ранее полученной информации в течение длительного времени. Процессы фиксации следов в долговременной памяти осуществляются лучше при повторных воздействиях, особенно биологически значимых раздражителей. Наиболее быстро процесс консолидации памяти происходит при действии эмоционально значимых раздражителей. Долговременная память по своему механизму качественно отличается от кратковременной памяти и не нарушается при таких экстремальных воздействиях на мозг, как механическая травма, электрошок, наркоз и др.
Механизм долговременной памяти окончательно не установлен. Несколько теорий с разных позиций объясняют механизмы долговременной памяти: морфологические теории, глиальная теория, медиаторная теория, молекулярные теории.
№ 9 Физиология сна. Структура сна: фазы сна и их проявления. Физиологические изменения во время сна. Современные представления о механизмах сна (П.К.Анохин).
Сон — физиологическое состояние, которое характеризуется потерей активных психических связей субъекта с окружающим миром.
Биологическое значение сна: активность мозга во время сна часто превосходит дневные уровни. Показано, например, что во время сна активность нейронов ряда структур мозга существенно возрастает. Во сне наблюдается и активация ряда вегетативных функций. Все это позволило рассматривать сон не как снижение процессов жизнедеятельности, а как активный физиологический процесс, активное состояние жизнедеятельности. Некоторые исследователи рассматривают сон наряду с бодрствованием как второе состояние жизнедеятельности организма.
Объективные признаки сна:
Потеря сознания. Сон прежде всего характеризуется потерей активного сознания субъекта, выключением его активных связей с окружающим миром. Глубоко спящий человек не реагирует на многие воздействия окружающей среды.
Фазовые изменения ВНД при переходе ко сну. Особенно отчетливо фазовые состояния наблюдаются при засыпании, т.е. при переходе от бодрствования ко сну.
Полиметрия. Наиболее объективно характеризуют состояние сна изменения электроэнцефалограммы и ряда вегетативных показателей. Для этого применяют методы полиметрии, когда на многоканальном полиграфе одновременно регистрируются ЭЭГ в нескольких отведениях и ряд физиологических функций: ЭКГ, дыхание, артериальное давление, температура тела, показатели газообмена.
Выделяют несколько стадий изменения ЭЭГ во время сна:
Медленноволновая фаза: для состояния бодрствования характерна низкоамплитудная высокочастотная ЭЭГ-активность. При закрывании испытуемым глаз и расслаблении «быстрая» бета-ЭЭГ-активность сменяется на более медленный альфа-ритм. В этот период происходит постепенное расслабление мышц, глаза закрываются, человек погружается в дремотное состояние; пробуждение в этой фазе происходит легко, достаточно слегка потревожить засыпающего. В течение следующего получаса на ЭЭГ альфа-волны начинают складываться в характерные «веретена». Стадия «веретен» примерно через 30 мин сменяется стадией высокоамплитудных медленных тета-волн. Пробуждение в эту фазу становится затруднительным. Снижаются ЧСС, кровяное давление, температура тела; сердцебиение и дыхание становятся регулярными. Стадия тета-волн сменяется стадией, когда на ЭЭГ нарастают высокоамплитудные сверхмедленные дельта-волны. Дельта – сон – период глубокого сна. Обычно при засыпании стадия медленноволнового сна занимает около 1—1,5 ч. Эта стадия сменяется появлением на ЭЭГ низкоамплитудной высокочастотной активности. Она получила по ЭЭГ-характеристике название парадоксального, или быстроволнового, сна. В состоянии быстроволнового сна испытуемые находятся в глубоком сне, их нельзя разбудить сильными раздражителями, но они просыпаются от малейшего шороха. Первое проявление парадоксального сна длится 6—10 мин. Затем на ЭЭГ снова возникают альфа-волны с последующими проявлениями фаз медленноволнового сна. Парадоксальный ЭЭГ-сон с интервалами 80—90 мин периодически сменяет медленноволновой сон.
Быстрые движения глаз. В соответствии с ЭЭГ-изменениями во время сна выявлены характерные быстрые движения глаз, которые точно совпадают с парадоксальной ЭЭГ-стадией сна. Установлено, что если спящего человека разбудить в фазу парадоксального сна, то он сообщает о снах. Этого не отмечается при пробуждении в фазу медленноволнового сна.
Корково – подкорковая теория Анохина: позволяет объяснить разные виды сна и его расстройства. Какова бы ни была причина сна, состояние сна связано с важнейшим механизмом – снижением восходящих активирующих влияний подкорковых образований на кору мозга.
№ ? Физиология мотиваций. Мотивация как компонент системной организации поведенческих актов. Виды мотиваций. Физиологические механизмы биологических мотиваций.
Мотивация — вызванное той или иной потребностью эмоционально окрашенное состояние организма, избирательно объединяющее нервные элементы различных уровней мозга. На основе мотиваций формируется поведение, ведущее к удовлетворению исходной потребности. Основные влечения организма представляют собой именно такую форму поведенческой деятельности, которая в отличие от рефлекторной деятельности как реакции на внешние стимулы, напротив, направлена на поиск специальных раздражителей внешней среды. К числу основных влечений, направленных на обладание определенными раздражающими предметами относятся, в первую очередь, влечение голода и ориентировочно – исследовательская деятельность.
Различают биологические и социальные мотивации:
Биологические мотивации, они же основные влечения, или низшие, простые, первичные мотивации. Биологические мотивации направлены на удовлетворение ведущих биологических потребностей индивидуумов по сохранению их вида или рода. К ним относятся мотивации голода, жажды, страха, агрессии, половые влечения, различные родительские, в частности материнские, температурные и другие влечения. Ведущими биологическими потребностями являются: 1) пищевая потребность, характеризующаяся уменьшением в организме уровня питательных веществ; 2) питьевая потребность, связанная с повышением осмотического давления; 3) температурная потребность — при изменении температуры тела; 4) половая потребность.
Всегда имеется ведущий параметр общей метаболической потребности — доминирующая потребность, наиболее важная для выживания особи или ее рода, которая строит поведенческий акт, направленный на ее удовлетворение.
Социальные мотивации, они же высшие, или вторичные, строятся на основе врожденных биологических мотиваций путем общения индивидуумов со средой обитания, родителями и окружающими их живыми существами, а у человека — и с социальной средой. В формировании социальных мотиваций значительное место принадлежит воздействию различных факторов внешней среды, обучению и, следовательно, механизмам памяти.
Мотивации как биологического, так и социального плана в целостной деятельности организма всегда являются компонентом системной архитектоники поведенческого акта. Мотивации играют существенную роль в организации таких ответственных системных механизмов, как стадия афферентного синтеза, принятия решения и предвидения потребного результата — акцептора результата действия. Не меньшее значение мотивация имеет в формировании эфферентной программы поведения — стадии эфферентного синтеза. Мотивации в своем становлении в развивающемся организме подлежат общим закономерностям развития функциональных систем — процессам системогенеза. В процессе онтогенетического развития отдельных функциональных систем мотивации, будучи их составным компонентом, созревают избирательно и последовательно.
Физиологические механизмы биологических мотиваций:
Биологические мотивации, будучи тесно связанными с метаболическими потребностями организма, строятся на основе врожденных, генетически детерминированных механизмов. Причиной биологических мотиваций являются раздражители внутренней среды, связанные с изменением различных показателей гомеостаза, определяющих нормальное течение обменных процессов в организме, например уровня питательных веществ, осмотического давления, состояния половых клеток, различных гормонов. Биологические мотивации могут активироваться или тормозиться специальными внешними «освобождающими» факторами внешней среды, например видом или запахом противника или полового партнера, пищи или других удовлетворяющих различные потребности раздражителей. Биологические мотивации нередко относятся к разряду сложных безусловных рефлексов, или инстинктов.