

Вопрос №1
IV.Взаимоотношения между организмами

1.Биотические связи
Среди
огромного разнообразия взаимосвязей
живых существ выделяют определенные
типы отношений, имеющие много общего у
 организмов
разных систематических групп.
организмов
разных систематических групп.
1.Симбиоз
Симбиоз1 - сожительство (от.греч.сим - вместе, биос - жизнь) - форма взаимоотношения, из которых оба партнера или хотя бы один извлекают пользу.
Симбиоз подразделяется на мутуализм, протокооперацию и комменсализм.
Мутуализм2 - форма симбиоза, при которой присутствие каждого из двух видов становится обязательным для обоих, каждый из сожителей получает относительно равную пользу, и партнеры (или один из них) не могут существовать друг без друга.
Типичный пример мутуализма - отношения термитов и жгутиковых простейших, обитающих в их кишечнике. Термиты питаются древесиной, однако у них нет ферментов для переваривания целлюлозы. Жгутиконосцы вырабатывают такие ферменты и переводят клетчатку в сахара. Без простейших - симбионтов - термиты погибают от голода. Сами же жгутиконосцы помимо благоприятного микроклимата получают в кишечнике пищу и условия для размножения.
Протокооперация3 - форма симбиоза, при которой совместное существование выгодно для обоих видов, но не обязательно для них. В этих случаях отсутствует связь именно этой, конкретной пары партнеров.
Примером протокооперации являются взаимоотношения мелких рыбок семейства губановых и крупных хищных мурен. Среди губановых имеются так называемые рыбы-чистильщики, освобождающие крупных рыб от наружных паразитов, находящихся на коже, в жаберной и ротовой полостях. Крупные хищники, в том числе мурены, страдающие от паразитов, приплывают в места обитания губанов и дают им возможность уничтожать паразитов даже у себя во рту, хотя могли бы с легкостью их проглотить.
Комменсализм - форма симбиоза, при которой один из сожительствующих видов получает какую-либо пользу, не принося другому виду ни вреда, ни пользы.
Комменсализм, в свою очередь, подразделяется на квартиранство, сотрапезничество, нахлебничество.
"Квартиранство"4 - форма комменсализма, при которой один вид использует другой (его тело или его жилище) в качестве убежища или своего жилья. Особую важность приобретает использование надежных убежищ для сохранения икры или молоди.
Пресноводный горчак откладывает икру в мантийную полость двухстворчатых моллюсков - беззубок. Отложенные икринки развиваются в идеальных условиях снабжения чистой водой.
"Сотрапезничество"5 - форма комменсализма, при которой несколько видов потребляют разные вещества или части одного и того же ресурса.
"Нахлебничество"6 - форма комменсализма, при которой один вид потребляет остатки пищи другого.
Примером перехода нахлебничества в более тесные отношения между видами служат взаимоотношения рыбы-прилипалы, обитающей в тропических и субтропических морях, с акулами и китообразными. Передний спинной плавник прилипалы преобразовался в присоску, с помощью которой та прочно удерживается на поверхности тела крупной рыбы. Биологический смысл прикрепления прилипал заключается в облегчении их передвижения и расселения.
2.Нейтрализм
Нейтрализм7 - тип биотической связи, при которой совместно обитающие на одной территории организмы не влияют друг на друга. При нейтрализме особи разных видов не связаны друг с другом непосредственно.
Например, белки и лось в одном лесу не контактируют друг с другом.
3.Антибиоз
Антибиоз - тип биотической связи, когда обе взаимодействующие популяции(или одна из них) испытывают отрицательное влияние друг друга.
Антибиоз подразделяется на аменсализм, хищничество, конкуренцию и паразитизм.
Аменсализм8 - форма антибиоза, при которой один из совместно обитающих видов угнетает другой, не получая от этого ни вреда, ни пользы.
Пример: светолюбивые травы, растущие под елью, страдают от сильного затемнения, в то время как сами на дерево никак не влияют.
Хищничество9 - тип антибиоза, при котором представители одного вида питаются представителями другого вида. Хищничество широко распространено в природе как среди животных, так и среди растений. Примеры: насекомоядные растения; лев, поедающий антилопу и т.д.
Кокуренция - тип биотических взаимоотношений, при котором организмы или виды соперничают друг с другом в потреблении одних и тех же обычно ограниченных ресурсов. Конкуренцию подразделяют на внутривидовую и межвидовую.
Внутривидовая кокуренция10 - соперничество за одни и те же ресурсы, происходящее между особями одного и того же вида. Это важный фактор саморегулирования популяции. Примеры: птицы одного вида конкурируют из-за места гнездования. Самцы многих видов млекопитающих (например, оленей) в период размножения вступают друг с другом в борьбу за возможность обзавестись семьей.
Межвидовая кокуренция11 - соперничество за одни и те же ресурсы, происходящее между особями разных видов. Примеры межвидовой кокуренции многочисленны. И волки, и лисы охотятся на зайцев. Поэтому между этими хищниками возникает конкуренция за пищу. Это не значит, что они непосредственно вступают в борьбу друг с другом, но успех одного означает неуспех другого.
Паразитизм12 - форма антибиоза, когда представители одного вида используют питательные вещества или ткани особей другого вида, а также его самого в
временного или постоянного местообитания.
Например, миноги нападают на треску, лососей, корюшку, осетров и других крупных рыб и даже на китов. Присосавшись к жертве минога питается соками ее тела в течение нескольких дней, даже недель. Многие рыбы погибают от нанесенных ею многочисленных ран.
Все перечисленные формы биологических связей между видами служат регуляторами численности животных и растений в сообществе, определяя его устойчивость.
Вопрос №2
МЕДИЦИНСКАЯ ПАРАЗИТОЛОГИЯ. ОБЩИЕ ВОПРОСЫ
1. ПРЕДМЕТ И ЗАДАЧИ
МЕДИЦИНСКОЙ ПАРАЗИТОЛОГИИ
Болезни животных и человека можно классифицировать по этиологическому
принципу как эндогенные и экзогенные. В основе эндогенных заболеваний
лежат аномалии структуры или функционирования наследственного аппарата.
Экзогенные заболевания имеют разную природу: это травмы, нарушения
питания, авитаминозы и т.д. Кроме того, это болезни, вызываемые живыми
организмами: вирусами, прокариотами и животными. Болезни, вызываемые
вирусами и прокариотическими организмами, называют инфекционными.
Болезни, вызываемые животными, называют инвазионными или паразитарными.
.
Медицинская паразитология изучает особенности строения и жизненных
циклов паразитов, взаимоотношения в системе паразит — хозяин, а также
методы диагностики, лечения и профилактики инвазионных болезней.
В связи с тем что большинство паразитов человека относится к типу
Простейшие Protozoa, а также к группе Черви (гельминты) — плоские
Plathelminthes и круглые Nemathelminthes, — в рамках паразитологии
выделяют разделы: медицинскую протозоологию и медицинскую
гельминтологию.
Немало животных, имеющих медицинское значение и в типе Членистоногие
Arthropoda. Некоторые из них сами являются возбудителями заболеваний,
другие — переносчиками возбудителей паразитарных и инфекционных
болезней. Биологию членистоногих — возбудителей и переносчиков (клещей и
насекомых) — изучает медицинская арахноэнтомология.
Паразиты могут обитать в любых органах человека, поэтому врач любой
специальности может встречаться с паразитарными заболеваниями и обязан
уметь распознавать их, лечить больных и проводить профилактику заражения
паразитами (рис. 18-1).
Рис. 1. Локализация паразитов в организме человека:
1 — головной мозг (эхинококк, цистицерки свиного цепня, трипаносома,
токсоплазма и др.), 2 — кровь (малярийный плазмодий, трипаносома и др;),
3 — легкие (эхинококк, легочный сосальщик и др.), 4 —.печень (многие
сосальщики, эхинококк, токсоплазма и др.), 5 — скелетные мышцы
(трихинелла, цистицерки свиного цепня и др.), 6 — подкожная клетчатка
(ришта и др.), 7 — мочеполовая система (шистосомы, трихомонада и др.), 8
— кожа открытых участков тела (лейщмания и др.), 9 — толстая кишка
(дизентерийная амеба, балантидий, власоглав и др.), 10 — тонкая кишка
(лямблия, все цепни, лентецы, аскарида, острица и др.)
Вопрос №3
Основные понятия паразитологии
По данным Всемирной организации здравоохранения из 50 млн. человек, ежегодно умирающих в мире, более чем у 16 млн. причиной смерти являются инфекционные и паразитарные заболевания. Этот класс болезней остаётся ведущим среди причин смертей человечества в настоящее время. По числу больных в мире, заражения кишечными гельминтозами занимают третье место. Паразитарные болезни являются причиной задержки психического и физического развития детей, снижают трудоспособность взрослого населения. Вызывая аллергизацию организма, поражённого человека, паразиты снижают сопротивляемость к инфекционным и соматическим заболеваниям, уменьшают эффективность вакцинопрофилактики. По оценке Всемирного банка экономический ущерб от кишечных гельминтозов занимает четвёртое место среди всех болезней и травм. Учитывая важность борьбы с паразитарными болезнями для многих стран, пятьдесят четвёртая сессия Всемирной ассамблеи здравоохранения в 2001 г. одобрила стратегию борьбы с гельминтозами до 2010г. Гельминты- паразитические черви из сборной группы плоских и первично-плоских червей. (Большой энциклопедический словарь (БЭС). Биология. 1998, стр.120.) Паразитизм- форма взаимоотношения двух различных организмов, принадлежащих к разным видам и носящая антагонистический характер, когда один из них (паразит) использует другого (хозяина) в качестве среды обитания или источника пищи. (БЭС. Биология. 1998, стр.448.) Глистные инвазии (гельминтозы)- заболевания, вызываемые паразитическими червями (гельминтами). В настоящее время известно около 300 гельминтозов человека, но широко распространены далеко не все. Почему же в наш развитый век, когда создаются сложнейшие компьютерные программы, строятся космические корабли, покоряются моря и океаны, люди не могут справиться с глистами.
Для этого существует несколько причин:
Эта болезнь из разряда «стыдно сказать»
К большинству гельминтов восприимчивость всеобщая
Низкая диагностическая ценность существующих методов исследования и, зачастую, неправильные подходы к терапии глистной инвазии
Паразитические черви- это уникальные живые существа. В процессе их эволюции, они приобрели возможность существовать за счёт хозяина. Многие глисты потеряли способность к самостоятельному передвижению, в значительной мере упростилась их нервная система. У глистов существуют органы, позволяющие паразиту удерживаться на одном месте – присоски и крючья. Многие глисты, обитающие в просвете кишечника, способны поглощать питательные вещества всей поверхностью тела. У паразитов изменилась сама форма, тело стало более утолщённым. Всем гельминтам присуща стадийность развития, т.е. на протяжении жизни гельминт проходит ряд последовательных стадий: чаще яйцо – личинка - взрослая особь. Гельминт проникает в организм человека на одной стадии развития, а покидает его на другой, что обусловливает практически незаразность больного. За редким исключением больной человек непосредственной опасности для окружающих не представляет, но является источником распространения гельминтов во внешнюю среду. Гельминты приобрели способность к длительному существованию в организме человека. Некоторые особи живут от 1,5 лет до 25 и более лет. И могут достичь гигантских размеров. Ещё одной особенностью гельминтов является их необычайная плодовитость. Например, взрослая самка аскариды способна отложить до 200 тысяч яиц в сутки
Паразитология (греч. <паразитос> - нахлебник, паразит, <логос> - учение) - комплексная наука, изучающая как самих паразитов, так и вызываемые ими болезни и методы борьбы с ними. Возникает вопрос: какие же взаимоотношения организмов называются паразитическими, как определить паразитизм? Паразитизм-ассоциация генетически разнородных организмов, основанная на пищевых связях и взаимообмене, при которой один (паразит) использует другого (хозяина) в качестве среды обитания и источника питания, причем оба партнера находятся в антагонистических отношениях различной степени остроты. Паразит вызывает в организме хозяина иммунобиологические реакции. Собственно паразитология состоит из протозоологии - науки о паразитических простейших и вызываемых ими болезнях; гельминтологии - изучающей паразитических червей и вызываемые ими болезни; арахнологии и энтомологии, которые изучают паукообразных и насекомых как возбудителей, так и переносчиков инфекционных и инвазионных болезней.
Виды паразитизма.
Животные организмы, ведущие паразитический образ жизни, принято делить на временных и стационарных.
Временные паразиты - организмы, которые совершают весь цикл развития от яйца до взрослой стадии вне организма хозяина (последнего они используют только для питания). Ими могут быть слепни, комары, постельные клопы и некоторые другие насекомые. Временные паразиты периодически нападают на животных (своих хозяев) с целью питания; они являются, как правило, эктопаразитами (наружными).
Стационарные паразиты инвазируют хозяина на продолжительное время (или в течение всей его жизни) не только для питания, но и для обитания. Обитают они на поверхности тела или в организме хозяина, а локализуются чаще всего во внутренних органах. Таким образом, стационарные паразиты являются в большей массе своей внутренними, или эндопаразитами. Подразделяются эти паразиты на постоянных и периодических. Постоянные паразиты (вши, чесоточные клещи и др.) обитают на хозяине или в его организме в течение всей своей жизни, совершая весь цикл биологического развития. Периодические паразиты инвазируют хозяина только на определенной стадии развития. Например, различные оводы поселяются в организме хозяина только в личиночной стадии, в то время как взрослые насекомые и куколки: паразитический образ жизни не ведут. Периодическими паразитами являются подавляющее большинство гельминтов.
К числу наиболее распространенных инвазионных болезней лошадей относят: пироплазмоз лошадей, случную болезнь, чесотку, микроспорию, различные энтомозы (гастрофилезы, симулидотоксикозы, куликоидесы), параскаридоз, оксиуроз, парафиляриоз, гематопиноз и др.
При анализе взаимодействия в системе “паразит-хозяин” разбираются пути инвазии и инфекции, в т.ч. трансмиссивный; при изучении “Развития паразитов” дается понятие о смене хозяев и факторах (генетической и негенетической природы), определяющих восприимчивость хозяина к тому или иному виду возбудителя.
+
http://www.my-coralclub.com/index.php?option=com_content&view=article&id=418:13&catid=79:3&Itemid=80
Вопрос №4
Популяции как хозяев, так и паразитов являются обязательными членами биогеоценозов, устойчивость которых зависит, в частности, от видового разнообразия живых организмов, входящих в их состав (см. § 16.1). Паразиты в экосистемах являются консументами второго и третьего порядков и играют существенную роль в биотическом круговороте веществ. Даже самые патогенные из них, вызывающие гибель большого числа особей хозяев, выступают, с одной стороны, как стабилизаторы численности хозяев, периодически изымая из популяций избыток организмов, который мог бы привести к нарушению экологического баланса. С другой стороны, наиболее тяжелое течение паразитарных заболеваний обычно наблюдается у особей с ослабленным иммунитетом, страдающих наследственными дефектами или с врожденной предрасположенностью к аллергическим реакциям. Гибель именно этих организмов оказывает на генетическую структуру популяций хозяина благотворную роль, элиминируя из его аллелофон-да аллели, снижающие жизнеспособность. Таким образом, взаимоотношения между популяциями хозяев и паразитов в условиях конкретных биогеоценозов способствуют их устойчивости и одновременно выступают как фактор естественного отбора, снижая неспецифический генетический груз популяции хозяина.
Кроме того, утрата биогеоценозами наиболее восприимчивых к заражению паразитами особей хозяина сопровождается и уничтожением части паразитарной популяции, гибнущей вместе с ним. Это обеспечивает активизацию микроэволюционных процессов в оставшейся части популяции паразитов, способствуя в конечном счете появлению у них новых адаптации.
В связи с социальностью человека в настоящее время паразитизм как фактор естественного отбора в человеческих популяциях значения практически не имеет. Однако целенаправленная борьба человека с паразитами, осуществляющаяся разными способами, несомненно является важным фактором эволюции самих паразитов.
Изучение распределения паразитов в популяциях хозяина показало, что оно зависит от многих факторов. Одним из них является возраст хозяина. Ряд паразитов чаще встречаются у взрослых хозяев. Действительно, у взрослых организмов больше шансов прийти в соприкосновение со многими паразитами и быть инвазированными. Например, влагалищная трихомонада поражает только половозрелых людей потому, что передается только половым путем.
Дифиллоботриозом и описторхозом человек заражается, поедая недостаточно термически обработанную рыбу. Такой путь заражения маловероятен для ребенка. Восточно-африканский трипаносомоз встречается чаще у людей среднего возраста — охотников, путешественников, участников геологоразведочных партий в необжитых саваннах Африки. Эта закономерность часто проявляется и у промежуточных хозяев: взрослые крупные рыбы имеют больше возможностей стать носителями метацеркарий сосальщиков или плероцеркоидов ленточных червей, чем мелкие молодые.
Другие паразиты чаще встречаются у детей. Причины этого разнообразны. Во-первых, это связано с незрелостью иммунной системы детей, а во-вторых, с не отработанными еще навыками личной гигиены. Поэтому для детей характерен особый спектр паразитарных заболеваний, в заражении которых большое значение имеет состояние иммунитета, а инвазия осуществляется просто при проглатывании цист, яиц или личинок, без участия промежуточных хозяев. Это лямблиоз, энтеробиоз, гименолепидоз, аскаридоз. Более частому заражению детей способствует их тесный и продолжительный контакт друг с другом в детских учреждениях.
На вероятность заражения также часто накладывает отпечаток профессия. Так, тениозом и тениаринхозом обычно заражаются работники мясокомбинатов, анкилостомидозами в умеренных широтах — шахтеры, а в тропиках — работники сельского хозяйства. Дифиллоботриозом чаще заражаются рыбаки, а альвеококкозом — охотники и лица, обрабатывающие меховое сырье.
Некоторые паразиты, в первую очередь простейшие, а также острица, угрица кишечная, карликовый цепень и трихинелла, попав в организм человека, способны размножаться в нем. То же касается личинок сосальщиков и некоторых ленточных червей в промежуточных хозяевах. Все это способствует тому, что в популяциях хозяина паразиты распределяются не равномерно, а перерассеянно. Это означает, что в меньшей части популяции хозяина сосредоточена большая часть популяции паразита.
Данная особенность имеет принципиальное значение для выживания хозяев, так как сводит к минимуму результаты массированных инвазий их паразитами, сохраняя основное ядро популяции интакт-ным. Перерассеянное распределение паразита в популяции хозяина необходимо учитывать при проведении профилактических мероприятий. Действительно, для выживания популяции паразита и передачи ее следующему поколению хозяев нередко важными оказываются лишь несколько особей или даже один организм, несущий в себе очень большое число паразитов.
ВЗАИМООТНОШЕНИЯ <ПАРАЗИТ - ХОЗЯИН>
(В. п. - х.) - один из вариантов вертикальных взаимоотношений организмов, при которых происходит передача вещества и энергии с одного трофического уровня на другой. Поскольку существуют суперпаразиты (т. е. <паразиты паразитов>, заключенные друг в друга наподобие матрешки, вплоть до четвертого порядка), то может формироваться особый вариант паразитарной пищевой цепи. Есть также примеры сложных В. п. - х. с посредником. Так, гетеротрофное растение-паразит подъельник паразитирует на грибах, разлагающих мертвое органическое вещество, но, кроме того, по гифам микоризного гриба как по шлангу выкачивает питательные элементы из корней ели. В естественных экосистемах В. п. - х. являются одним из важных факторов поддержания экологического равновесия, причем в процессе длительной коэволюции паразитов и хозяев вырабатываются специальные механизмы, которые позволяют им устойчиво сосуществовать. У хозяев вырабатывается целый ряд защитных реакций, главные из которых: - иммунный ответ организма, т. е. возникновение биохимических реакций, которые сдерживают массовое развитие паразитов; - сбрасывание зараженных частей (это особенно характерно для растений-хозяев, которые сбрасывают сильно зараженные листья). В этом случае паразиты продолжают жить уже как детритофаги; - выработка устойчивости к влиянию паразитов за счет быстрого роста здоровых тканей взамен пораженных (это имеет место при поедании тканей растений тлями); - изоляция органов поражения как <зеленых островов> (формирование галлов у дуба, орешника и других растений после того, как насекомое-паразитоид отложит в ткани листа яйцо). В этом случае ответ запрограммирован: в генной памяти хозяина записана реакция на поселение паразитоида; - уменьшение плотности популяций хозяев, что снижает вероятность распространения паразита и заражения им. Зараженные животные менее подвижны и становятся более легкой добычей хищников, таким образом снижая долю зараженных особей в популяции; - формирование гетерогенных популяций хозяев, в составе которых есть экотипы, устойчивые к паразитам. Эти экотипы являются основой адаптивной селекции на повышение устойчивости культурных растений к грибковым заболеваниям. Для естественных экосистем формирование экологического равновесия между популяциями паразитов и их хозяев - нормальное явление. В силу того, что паразиты связаны с ограниченным кругом хозяев, эта связь математически описывается много проще, чем связь между хищниками и их жертвами. Во многих случаях проявляется закономерность: плотность популяций обоих видов изменяется циклически, но пики плотности паразитов запаздывают по отношению к пикам плотности хозяев. Ситуация изменяется в антропогенных экосистемах (особенно в сельскохозяйственных), где заражение паразитами может привести к существенному падежу скота. Представляют опасность взаимоотношения паразитов и человека, который может заболевать гельминтозами, вызываемыми разными видами глистов, лямблиозом, болезнями бактериальной и вирусной природы. Катастрофическими были последствия заноса паразитов в новые районы, где у их потенциальных хозяев отсутствуют механизмы снижения отрицательных эффектов влияния паразитов. Уже в ХХ столетии произошли ботанические катастрофы в Америке (гибель зубчатого каштана от занесенного туда из Китая паразитического гриба, вызывающего <рак каштана>) и Европе, где от <голландской болезни> почти полностью исчез вяз. Болезнь вызывает гриб Ophiostoma ulmi, который переносится жуком-короедом. К настоящему времени американские генетики получили устойчивые к паразиту экотипы вяза, разработана специальная методика <лечения> больных деревьев. Очевидно, что невозможно восстановить каштановые леса, но каштан может снова стать украшением парков.
Воздействие паразитов на хозяина:
1. Механическое воздействие ( механическое давление на ткани,
воздействие органов прикрепления, закупорка кровеносных
сосудов, кишечника и т.д.)
2. Паразитарная кастрация.
3. Отнятие пищи.
4. Нарушение обменных процессов.
5. Роль паразитов в открытии ворот для вторичной инфекции
Реакции хозяина на поселение паразита.
1. Клеточные и тканевые реакции
2. Гуморальные реакции
3. Приобретенный иммунитет
Специфичность паразитов. Характеристика явления специфичности.
Специфичность и встречаемость. Специфичность на разных стадиях
развития паразитов. Специфичность по отношению к промежуточным и
резервуарным хозяевам. Факторы, обуславливающие специфичность.
Возникновение специфичности. Специфичность как историческая
категория. Условия становления системы паразит – хозяин по Павловскому.
Вопрос №5
АДАПТАЦИИ К ПАРАЗИТИЧЕСКОМУ ОБРАЗУ ЖИЗНИ. ОСНОВНЫЕ ТЕНДЕНЦИИ
Переход к паразитическому образу жизни сопровождается появлением у паразитов ряда адаптации, облегчающих их существование, развитие и размножение в специфических условиях организма хозяина Разнообразие форм паразитизма, различное систематическое положение паразитов (их принадлежность к разным отрядам, классам и типам), а также обитание их в разных органах и системах хозяина обусловливают многообразие этих адаптации.
Однако некоторые приспособления являются абсолютно универсальными. К ним в первую очередь относятся высокая плодовитость и особенности половой системы. Действительно, возможность оставления потомства и попадания его в благоприятную среду — организм хозяина — у паразитов часто ничтожна. В связи с этим интенсивность размножения паразитов по сравнению со свободноживущими формами гораздо более велика. Достигается это разными способами. У многоклеточных это сильная степень развития половой системы и образование огромного количества половых продуктов. Этому способствуют первичный гермафродитизм плоских червей, изначально высокая плодовитость круглых червей и основной массы членистоногих. Нередко высокая интенсивность полового размножения дополняется размножением личиночных стадий жизненного цикла. Особенно это характерно для сосальщиков, личинки которых размножаются партеногенетически, а у некоторых ленточных червей — внутренним или наружным почкованием.
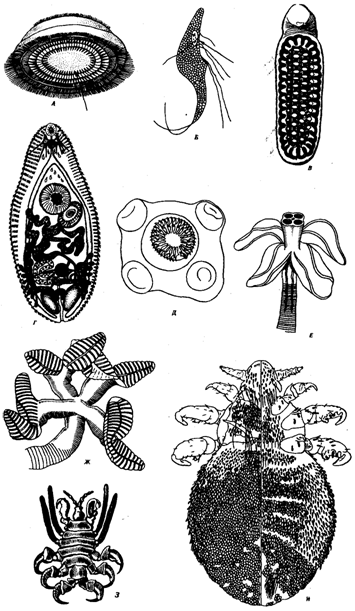
Рис. 18.4. Органы прикрепления паразитов
Присасывательные диски у инфузории (Л) и лямблии (Б) (указаны стрелками), сложная присоска (В) и кутикулярные шипики на коже (Г) у сосальщиков, крючья и сложные присоски на головках ленточных червей (Д, Е, Ж), конечности, служащие для прикрепления, у ракообразного (3) и насекомого (И)
Некоторые паразиты из типа простейших приобретают способность к множественному делению — шизогонии, когда из одного паразита может образоваться более 1000 дочерних особей, или к спорогонии, в результате которой из одной особи могут образоваться десятки тысяч организмов следующего поколения. Практически у всех эктопаразитов и паразитов, обитающих в полостных органах, имеются адаптации для прикрепления к телу хозяина. Они встречаются у простейших (присасывательные диски лямблии), у гельминтов (присоски, шипики, крючья плоских червей, хитинизированный ротовой аппарат ряда круглых червей) и паразитических членистоногих (своеобразные конечности) (рис. 18.4).
Эндопаразиты, обитающие в полостных органах, имеют покровы, обладающие антиферментными свойствами, быстро регенерирующие либо вообще непроницаемые для ферментов хозяина. Паразиты, живущие в тканях, часто там инкапсулируются (рис. 18.5).
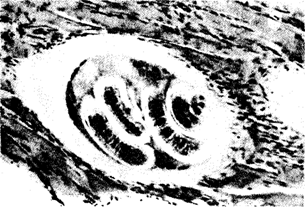
Рис. 18.5. Личинка трихинеллы, инкапсулированная в мышце
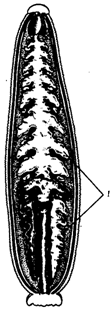
Рис. 18.6. Пищеварительная система пиявки:
1 – слепые выросты кишечника
Паразиты, питающиеся кровью (представлены в основном членистоногими), имеют колюще-сосущий ротовой аппарат, а также сильно растяжимый хитиновый покров, часто разветвленную пищеварительную трубку (рис. 18.6), антикоагулянтные свойства слюны и консервантные свойства ферментов пищеварительной системы.
Эндопаразиты, активно отыскивающие хозяина, обладают органами ориентации в среде, используемыми для поисков хозяина (светочувствительные глазки, термо- и хеморецепторы), и органами передвижения (рис. 18.7).
Передний конец тела паразитов, внедряющихся в организм хозяина, снабжен органами проникновения — специализированными железами, колющими стилетами и т.д. Это касается даже некоторых простейших, способных проникать в ткани хозяина через неповрежденные покровы.
Все паразиты, развивающиеся со сменой хозяев, используют в качестве таковых виды, связанные между собой непосредственными пищевыми взаимоотношениями или обитающие с ними в одной среде. Большинство промежуточных хозяев являются источником питания для основных. Другой распространенный путь попадания паразита в организм хозяина — это использование многочисленных переносчиков, которые обеспечивают не только постоянную циркуляцию паразитов в экологических системах, но и их широкое расселение.
Высшей степенью адаптации паразитов к хозяевам является наблюдаемая часто полная зависимость паразита от жизнедеятельности хозяев. При этом паразит нередко вызывает такие реакции хозяина, которые обеспечивают максимальную вероятность заражения последнего. Так, самки остриц, откладывая яйца в области анального отверстия, вызывают зуд. Расчесывание зудящих мест способствует распространению яиц этого паразита руками по окружающим предметам. Таким же образом обеспечивается расселение чесоточного клеща. Зуд в пораженной конечности, прекращающийся от соприкосновения с водой, способствует циркуляции в природе такого паразита, как ришта. Высокая температура больных паразитарными заболеваниями, распространяющимися с помощью кровососущих членистоногих, привлекает переносчиков нередко с больших расстояний и также оказывается полезной для паразитов.
Нередко особенности жизнедеятельности паразитов оказываются синхронизированными с образом жизни хозяев. Так, откладка яиц шистосомами происходит обычно в самое жаркое время суток, когда наиболее вероятным оказывается контакт хозяев с водой, куда для развития должны попасть яйца этих паразитов. В это же время в поверхностных слоях воды скапливаются в поисках хозяев церкарии этих шистосом. Таким образом облегчается циркуляция паразита сразу на двух стадиях его жизненного цикла (см. разд. 20.1.1.2). Если в циркуляцию паразита включены несколько хозяев, то наблюдаются их взаимные адаптации, оказывающиеся выгодными паразитам и обеспечивающие его эффективное развитие. Так, выход микрофилярий в кровеносные сосуды человека происходит в часы суток, соответствующие периоду максимальной активности кровососущих насекомых, являющихся их переносчиками (см. разд. 20.2.1.2).
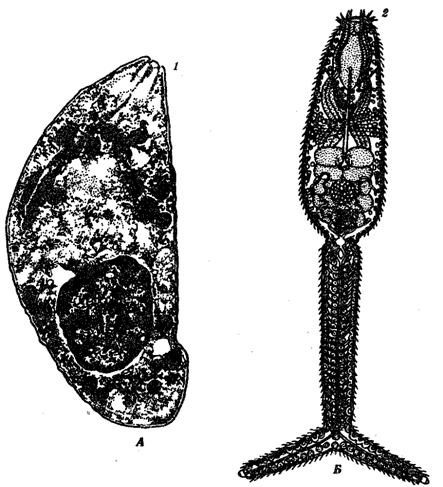
Рис. 18.7. Приспособления, служащие для проникновения в организм хозяина. А— простейшее токсоплазма; Б — церкарии — личинка кровяного сосальщика:
1 — коноид, 2 — колющие стилеты
Нередко паразиты даже модифицируют поведение одних хозяев таким образом, что в результате облегчается их попадание к другим. Так, рыбы, пораженные личинками ленточных червей, плавают в основном у поверхности воды и чаще вылавливаются рыбаками и хищными животными. Ленточные черви, использующие в качестве промежуточных хозяев копытных животных, снижают их жизнеспособность, и, таким образом, хищники поедают их в первую очередь. Сосальщики, заражающие травоядных животных и человека через случайное проглатывание насекомых, вызывают обездвиживание последних и плотное прикрепление к растениям, облегчающее им попадание в пищеварительный тракт хозяина (см. раздел 20.1.1.3. стр. 245).
Одновременно с перечисленными признаками свойства паразитов переживать неблагоприятные условия внешней среды являются также несомненными адаптациями к паразитизму. Большинство простейших, заражение которыми происходит без участия переносчиков, во внешней среде способны инцистироваться. Яйца большинства гельминтов обладают феноменальной устойчивостью к неблагоприятным воздействиям. Капсулы с личинками трихинелл переносят не только промораживание, кипячение, но и многократное прохождение через пищеварительную систему рыб, земноводных, птиц, насекомых и ракообразных, не теряя жизнеспособности.
Для большинства паразитов, переживших нахождение во внешней среде или в промежуточном хозяине в покоящихся стадиях и попавших в организм окончательного хозяина, существует комплекс условий, являющийся сигналом к началу активной жизнедеятельности. У млекопитающих он часто неспецифичен: это температура тела около 37°С, водная среда и высокая ее кислотность, а также наличие ферментов желудочного сока. Такие условия характерны для желудка любого млекопитающего, поэтому цисты, яйца и другие инвазионные стадии паразитов, обитающие у разных млекопитающих могут, попадая в желудок человека, начинать развитие. Некоторые из них проходят лишь часть цикла и, не находя специфических условий, гибнут (см. разд. 20.2.1.3), успевая, однако, привести к тяжелым последствиям.
Указанные особенности паразитов, общие для многих из них, не состоящих в родстве, возникли в разных их группах независимо друг от друга, иллюстрируя конвергентный характер эволюции организмов разных видов, классов и типов, адаптирующихся к сходным условиям.
Изменение формы тела у эктопаразитов: сплющивание тела в дорзо –
вентральном направлении, способствующее лучшему прикреплению к телу
хозяина; укорачивание тела; сглаживание метамерии у сегментированных
видов; редукция конечностей,
Изменение формы тела у эндопаразитов: кишечные эндопаразиты –
тенденция к удлинению тела, расчленение тела в поперечном направлении,
сплющивание тела; внутренностные эндопаразиты – округлая форма тела,
тенденция к увеличению поверхности тела, многократное ветвление одного
из концов тела.
Общего для всех паразитов влияния образа жизни их на размеры не
наблюдается. Но в тех случаях, когда такое влияние есть, оно сказывается
в увеличении размеров тела при переходе к паразитическому образу жизни,
Органы прикрепления составляют одну из самых характерных
особенностей большинства паразитов. Несмотря на необычайно широкое
распространение органов прикрепления в животном мире, устроены они
сравнительно однообразно, что дает повод к развитию многочисленных
конвергенций. В основном существует два типа прикрепительных аппаратов
– крючки ( выросты, различающиеся по форме и расположению) и присоски
(более или менее глубокие ямки на теле, обведенные валиком особо
дифференцированной мускулатуры). В отдельных случаях используются
другие способы прикрепления: клещи (принцип захлопывания двух
створок), тип обхватывающего крепления (выросты тела паразита
обхватывают участок тела в виде кольца, без ущемления), распорки,
стилеты, стрекательные нити, липкие или прядильные нити, нитевидные
отростки тела, стебельки.
Адаптивные изменения в пищеварительной системе. Эволюция и
усовершенствование строения ротовых аппаратов. Появление у
гематофагов специальных органов повреждения покровов хозяина,
приспособления к увеличению емкости кишечника. Тенденция к редукции и
исчезновению кишечника у различных групп эндопаразитов. Исчезновение
кишечника в онтогенезе отдельных видов.
Адаптации половой системы к паразитизму. Функция размножения у
большинства паразитов становится доминирующей; кроме обычных
способов размножения, часто появляются еще иные, усиливающие
способность животного к размножению. Паразитический образ жизни,
прикованность к организму хозяина затрудняет встречу между особями
одного пола. Следствием этого − выраженная у паразитических, а особенно
у эндопаразитических животных, тенденция к выработке
гермафродитизма. Первичный (происхождение от гермафродитных
свободных предков) и вторичный гермафродитизм (возникновение
гермафродитизма под влиянием паразитического образа жизни) паразитов.
Адаптации к перекрестному оплодотворению как путь к вторичному
разделению полов.
Помимо тенденции к гермафродитизму для паразитов характерна
громадная плодовитость, огромное число продуцируемых яиц. Это
свойство, называемое законом большого числа яиц, служит
приспособлением, помогающим паразитам в борьбе с множеством
неблагоприятных условий, которые приходится преодолевать во время
сложного жизненного цикла. Увеличение половой продукции влечет за
собой ряд морфологических изменений, состоящих главным образом в
увеличении размеров половых органов, умножении числа половых
комплексов. К другим приспособлениям для умножения числа особей и
продления жизни вида можно отнести: протогенез и неотению,
партеногенез, включение в жизненный цикл процессов бесполого
размножения, удлинение репродуктивного цикла с целью удлинения сроков
репродуктивной деятельности или, наоборот сокращение цикла с целью
увеличения числа поколений, создание мощных и устойчивых оболочек,
защищающих покоящиеся стадии от агрессивных факторов внешней среды.
Адаптации нервной системы к паразитизму. У всех паразитов, а в
особенности у эндопаразитов, нервная система более или менее
упрощается.
Тема 3. Физиологические адаптации к паразитизму.
Размножение паразитов. Переход от амфигенного к
партеногенетическому размножению. Включение в цикл развития
бесполого размножения. Полиэмбриония.
Адаптации паразитов к дыханию. Полостные и тканевые паразиты
находятся в таких же условиях дыхания, как и соседние ткани самого
хозяина, т.е. кислород доставляется им кровью хозяина или
непосредственно его органами дыхания. Кишечные эндопаразиты живут в
почти бескислородной среде и обладают поэтому анаэробным дыханием.
Главным источником энергии служит гликоген, продукты анаэробного
расщепления которого представлены жирными кислотами (валериановая
кислота). Эти вредные для организма вещества перестраиваются в
нейтральные жиры, которые откладываются в тканях или выносятся наружу.
Это так называемый «экскреторный жир», который не может быть
использован в качестве источника энергии из – за дефицита кислорода.
Процессы анаэробного расщепления крайне невыгодны в энергетическом
отношении. Поэтому в тканях гельминтов накапливаются запасы гликогена,
расходуемого в огромных количествах.
Соотношение аэробных и анаэробных процессов у разных видов
эндопаразитов и факторы, обуславливающие это соотношение. Условия
дыхания у внутриклеточных паразитов.
Адаптация эндопаразитов к улавливанию кислорода из тканей
хозяина. Смена аэробных и анаэробных фаз у паразитов со сложным
циклом развития.
Адаптации паразитов к питанию. Переход паразитов от питания
частью покровов к питанию секретом желез и к гематофагии. Случайная и
облигатная гематофагия эктопаразитов.
Адаптации к питанию у эндопаразитов. Направление в эволюции в
характере питания эндопаразитов – переход от обычного способа питания к
специфически паразитическому способу питания..
Вопрос №6
Систематика живых организмов. Таксономия. Номенклатура.
Систематика живых организмов — одна из наиболее сложных задач биологии. Систематика концентрирует все основные достижения науки — чем они более конкретны, тем более точна классификация. Любая классификация живых организмов призвана показать степень сходства и предполагаемой эволюционной взаимосвязи (при этом более высокие категории — ёмкие и широкие, а более низкие — конкретны и ограничены). Отличительные признаки в строении растений и животных, служившие до середины XIX века основой систематики живых существ, видны с первого взгляда.
В первую очередь, эти различия вытекают из способов питания. Животные поглощают органические вещества, перевариваемые и всасываемые в пищеварительном тракте. В упрощённом виде развитие животного мира должно обеспечивать создание всасывающих внутренних поверхностей, кои у всех и существуют — от кишечнополостных до млекопитающих. Растения имеют принципиально иное строение, обусловленное в первую очередь иным типом питания. Минеральные вещества они всасывают из почвы корневой системой, а органические — образуют сами в процессе фотосинтеза.
Принципы классификации изучает особый раздел систематики — таксономия [от греч. taxis, расположение, порядок, + nomos, закон]. Все существующие классификации форм жизни весьма разнородны, ни одна из них не является полной, всеобъемлющей и принятой повсеместно. Чёткие границы мира растений и мира животных рухнули после открытия микроорганизмов. Для этого — третьего — царства живых существ Эрнст Хёккель (1866) предложил собирательное название протисты [от греч. protistos, первейший]. Всех их отличает более простое, чем у животных и растений, строение клетки. Выcшие протисты (грибы, водоросли и простейшие) — эукариоты [от греч. еu- хороший, добротный + karyon, ядро] — имеют морфологически обособленное ядро и митотически делятся, чем весьма напоминают растительные и животные клетки. Более просто организованную группу составляют прокариоты [от греч. pro-, предшествующий + karyon, ядро] — бактерии и сине-зелёные водоросли, чьи клетки не имеют мембраны вокруг вещества ядра. Позднее представителей микромира дополнили неклеточные формы жизни — вирусы, плазмиды, вироиды и др.
Биологи́ческая система́тика — научная дисциплина, в задачи которой входит разработка принципов классификации живых организмов и практическое приложение этих принципов к построению системы. Под классификацией здесь понимается описание и размещение в системе всех существующих и вымерших организмов[1].
Завершающим этапом работы систематика, отражающим его представления о некой группе живых организмов, является создание Естественной Системы. Предполагается, что эта система, с одной стороны лежит в основе природных явлений, с другой стороны является лишь этапом на пути научного исследования. В соответствии с принципом познавательной неисчерпаемости природы естественная система недостижима[2].
«Углублённое изучение уже известных групп, всё более разъясняя их взаимные соотношения, будет требовать других сопоставлений или, точнее сказать, перестановки членов. Нам кажется, что естественная система всегда будет подвергаться постоянным изменениям, так как каждая попытка может быть выполнена только в связи с состоянием научных знаний своего времени.»
— К. М. Бэр[3]
Основные цели систематики:
наименование (в том числе и описание) таксонов,
диагностика (определение, то есть нахождение места в системе),
экстраполяция, то есть предсказание признаков объекта, основывающееся на том, что он относится к тому или иному таксону. Например, если на основании строения зубов мы отнесли животное к отряду грызунов, то можем предполагать, что у него имеется длинная слепая кишка и стопоходящие конечности, даже если нам неизвестны эти части тела.
Систематика всегда предполагает, что:
окружающее нас разнообразие живых организмов имеет определённую внутреннюю структуру,
эта структура организована иерархически, то есть разные таксоны последовательно подчинены друг другу,
эта структура познаваема до конца, а значит, возможно построение полной и всеобъемлющей системы органического мира («естественной системы»).
Эти предположения, лежащие в основе любой таксономической работы, можно назвать аксиомами систематики[1].
Современные классификации живых организмов построены по иерархическому принципу. Различные уровни иерархии (ранги) имеют собственные названия (от высших к низшим): царство,тип или отдел, класс, отряд или порядок, семейство, род и, собственно, вид. Виды состоят уже из отдельных особей.
Принято, что любой конкретный организм должен последовательно принадлежать ко всем семи категориям. В сложных системах часто выделяют дополнительные категории, например, используя для этого приставки над- и под- (надкласс, подтип и т. п.). Каждый таксон должен иметь определённый ранг, то есть относиться к какой-либо таксономической категории.
Этот принцип построения системы получил название Линнеевской иерархии, по имени шведского натуралиста Карла Линнея, труды которого были положены в основу традиции современной научной систематики.
Сравнительно новым является понятие надцарства, или биологического домена. Оно было предложено в 1990 Карлом Вёзе и ввело разделение всей биомассы Земли на три домена: 1)эукариоты (домен, объединивший все организмы, клетки которых содержат ядро); 2) бактерии; 3) археи.
[править]История систематики
Первые известные нам попытки классифицировать формы жизни предприняли в античном мире Гептадор, а затем Аристотель и его ученик Теофраст, которые объединяли всё живое в соответствии со своими философскими взглядами. Они дали довольно подробную систему живых организмов. Растения были разделены ими на деревья и травы, а животные — на группы с «горячей» и «холодной» кровью. Последний признак имел большое значение для выявления собственной, внутренней упорядоченности живой природы. Так родилась естественная система, отражающая упорядоченность, имеющуюся в природе.[1]
В 1172 году арабский философ Аверроэс сделал сокращённый перевод трудов Аристотеля на арабский язык. Его собственные комментарии были утеряны, но сам перевод дошёл до наших дней на латыни.
Большой вклад сделал швейцарский профессор Конрад Геснер (1516—1565).
Эпоха великих открытий позволила учёным существенно расширить знания о живой природе. В конце XVI — начале XVII веков начинается кропотливое изучение живого мира, вначале направленное на хорошо знакомые типы, постепенно расширившееся, пока, наконец, не сформировался достаточный объём знаний, составивший основу научной классификации. Использование этих знаний для классификации форм жизни стало долгом для многих известных медиков, таких как Иероним Фабриций (1537—1619), ученик Парацельса Северинус (1580—1656), естествоиспытатель Уильям Гарвей (1578—1657), английский анатом Эдвард Тайсон (1649—1708). Свой вклад сделали энтомологи и первые микроскописты Марчелло Мальпиги (1628—1694), Ян Сваммердам (1637—1680) и Роберт Гук (1635—1702).
Английский натуралист Джон Рей (1627—1705) опубликовал важные работы по растениям, животным и натуральной теологии. Подход, использованный им при классификации растений в его «Historia Plantarum», стал важным шагом по направлению к современной таксономии. Рей отверг дихотомическое деление, которое использовалось для классификации видов и типов, предложив систематизировать их по схожести и отличиям, выявленным в процессе изучения.
[править]Линней
Порядок есть подразделение классов, вводимое для того, чтобы не разграничивать роды в числе большем, чем их легко может воспринять разум. Карл Линней |
Через два года после смерти Джона Рея родился Карл Линней (1707—1778), чья работа «Systema Naturae» (1735) была переиздана по меньшей мере тринадцать раз ещё при его жизни. Он разделил природный мир на три царства: минеральное, растительное и животное. Линней использовал четыре уровня (ранга): классы, отряды, роды и виды.
Линней определил основные положения научной систематики. Главным в систематике, по мнению Линнея, является построение естественной системы, которая, в отличие от каталожного списка, «сама по себе указывает даже на пропущенные растения». Он же был автором одной из популярных искусственных систем растений, в которой цветковые растения распределялись по классам в зависимости от числа тычинок в цветке[1].
Введённый Линнеем метод формирования научного названия для каждого из видов используется до сих пор (применявшиеся ранее длинные названия, состоящие из большого количества слов, давали описание видов, но не были строго формализованы). Использование латинского названия из двух слов — название рода, затем видовой эпитет — позволило отделить номенклатуру от таксономии. Данное соглашение о названиях видов получило имя «бинарная номенклатура».
[править]После Линнея
В конце XVIII века Антуан Жюссьё ввёл категорию семейства, а в начале XIX века Жорж Кювье сформулировал понятие о типе животных. Вслед за этим категория, аналогичная типу, —отдел — была введена для растений.
Чарлз Дарвин предложил понимать естественную систему как результат исторического развития живой природы. Он писал в книге «Происхождение видов»:
…общность происхождения <…> и есть та связь между организмами, которая раскрывается перед нами при помощи наших классификаций.
Это высказывание положило начало новой эпохе в истории систематики, эпохе филогенетической (то есть основанной на родстве организмов) систематики.[1]
Дарвин предположил, что наблюдаемая таксономическая структура, в частности, иерархия таксонов, связана с их происхождением друг от друга. Так возникла эволюционная систематика, ставящая во главу угла выяснение происхождения организмов, для чего используются как морфологические, так и эмбриологические и палеонтологические методы.
Новый шаг в этом направлении был сделан последователем Дарвина, немецким биологом Эрнстом Геккелем. Из генеалогии Геккель заимствовал понятие «генеалогическое (родословное) древо». Родословное древо Геккеля включало все известные к тому времени крупные группы живых организмов, а также некоторые неизвестные (гипотетические) группы, которые играли роль «неизвестного предка» и помешались в развилках ветвей или в основании этого древа. Такое чрезвычайно наглядное изображение очень помогло эволюционистам, и с тех пор — с конца XIX века — филогенетическая систематика Дарвина—Геккеля господствует в биологической науке. Одним из первых следствий победы филогенетики стало изменение последовательности в преподавании курсов ботаники и зоологии в школах и университетах: если раньше изложение начинали с млекопитающих (как в «Жизни животных» А. Брема), а затем спускались «вниз» по «лестнице природы», то теперь изложение начинают с бактерий или одноклеточных животных.[1]
Геккель очень хотел, чтобы на каждой развилке дерева можно было разместить какой-нибудь организм. Такой организм и был бы родительской (предковой) формой для всей ветки. Но если такие организмы и находили, впоследствии признавали их не предками, а «боковыми ветвями» эволюции. Так произошло, например, с тупайями, археоптериксом, ланцетником,трихоплаксом и многими другими организмами. Геккель мечтал найти организм, который можно было бы поместить в самое основание дерева, и даже однажды сообщил, что он найден. Организм представлял собой комок слизи и получил название батидий, но вскоре оказалось, что это — продукт деградации морских животных. Такое существо (по-английски оно называется last common ancestor, сокращённо LCA) не найдено до сих пор[1].
[править]Наименование и описание таксонов
К началу XX века в систематике оформилось семь основных таксономических категорий:
царство — regnum
тип — phylum (у растений отдел — divisio)
класс — classis
отряд (у растений порядок) — ordo
семейство — familia
род — genus
вид — species
Любое растение или животное должно последовательно принадлежать ко всем семи категориям. Часто систематики выделяют дополнительные категории, используя для этого приставкипод- (sub-), инфра- (infra-) и над- (super-), например: подтип, инфракласс, надкласс. Такие категории обязательными не являются, то есть при систематизации объекта их можно пропустить. Кроме того, часто выделяются и другие категории: раздел (divisio) между подцарством и надтипом у животных, когорта (cohors) между подклассом и надпорядком, триба (tribus) междуподсемейством и родом, секция (sectio) между подродом и видом, и так далее. Часто такие категории используются лишь в систематике каких-то конкретных таксонов (например, насекомых).
Для того чтобы избежать синонимии (то есть разных названий одного и того же таксона) и омонимии (то есть одного названия для разных таксонов), в настоящее время номенклатура регулируется номенклатурными кодексами, позволяющими деление на уровни (см. Ранг (биологическая систематика)),— отдельно для растений, животных и микроорганизмов. Во всех номенклатурных кодексах используются три основные принципа номенклатуры: приоритета, действительного обнародования и номенклатурного типа. Кроме того, названия всех таксонов должны даваться по-латыни (от латинских и греческих корней либо от личных имён или народных названий), а название вида должно быть бинарным, то есть состоять из названия рода и видового эпитета. Например, латинское название картофеля — Solanum tuberosum L. (последнее слово обозначает автора названия — в данном случае это Карл Линней; в зоологии часто ставят ещё и год действительного обнародования).
Каждый таксон обязательно должен иметь ранг, то есть относиться к какой-либо из перечисленных категорий. Таким образом, ранг — это мера соответствия таксонов друг другу; например, семейство Капустные и семейство Кошачьи — сопоставимые категории. Нет, однако, общепринятого способа вычисления ранга, и поэтому разные систематики выделяют ранги по-разному.[1]
[править]Диагностика таксонов
Под диагностикой понимают прежде всего составление таблиц для определения организмов (так называемых определительных ключей). Со времён Ж. Б. Ламарка наибольшее распространение получили дихотомические ключи, в которых каждый пункт (ступень) разделён на тезу и антитезу, снабжённые указаниями о том, к какой ступени нужно перейти дальше. Сейчас почти вся флора и фауна Земного шара охвачена определительными ключами.
В практической работе биолог-систематик руководствуется несколькими основными принципами и приёмами. Во-первых, классификация должна быть разбиением, то есть никакой таксон не может относиться сразу к двум группам одинакового ранга, и наоборот, каждый таксон должен относиться к какому-либо надтаксону (не должно быть неклассифицированного «остатка»). Во-вторых, классификация должна производиться по одному основанию, то есть признаки, используемые для классификации, должны быть альтернативными (нельзя делить на «растения с цветками» и «древесные растения»). В-третьих, классификация должна производиться по значимым признакам (например, нельзя использовать признаки роста и веса). В-четвертых, классификация должна проводиться по максимальному числу признаков (взятых из самых различных областей биологии — от морфологии до биохимии). Начинают классификацию с определения границ исходного таксона, затем выделяют элементарные таксоны (например, виды), подлежащие классификации. На следующем этапе происходит группировка таксонов. Иногда эту процедуру приходится повторять, покуда не будет достигнут приемлемый результат. Разные направления систематики различаются прежде всего методами группировки.[1]
[править]Иерархия
Надцарство
Царство
Подцарство
Надтип/Надотдел
Тип/Отдел
Клада
Подтип/Подотдел
Надкласс
Класс
Подкласс
Инфракласс
Надотряд/Надпорядок
Отряд/порядок
Подотряд/Подпорядок
Инфраотряд
Надсемейство
Семейство
Подсемейство
Надтриба
Триба
Подтриба
Род
Подрод
Надсекция
Секция
Подсекция
Ряд
Подряд
Вид
Подвид
Вариетет/Разновидность
Подразновидность
Форма
Подформа
[править]Современные разработки
В настоящее время принято, чтобы классификация там, где это допустимо, следовала принципам эволюционизма.
Обычно биологические системы создаются в виде списка, в котором каждая строчка соответствует какому-нибудь таксону (группе организмов). С 1960-х развивается направление систематики, называемое «кладистика» (или филогенетическая систематика), которое занимается упорядочиванием таксонов в эволюционное дерево — кладограмму, то есть схему взаимоотношений таксонов. Если таксон включает всех потомков некой предковой формы, он является монофилетическим. В. Хенниг формализовал процедуру выяснения предкового таксона, и в своей кладистической систематике положил в основу классификации кладограмму, строящуюся при помощи компьютерных методик. Это направление является ныне ведущим в странах Европы и США, особенно в сфере геносистематики (сравнительного анализа ДНК и РНК)[1].
Р. Сокэл и П. Снит в 1963 году основали так называемую численную (нумерическую) систематику, в которой сходство между таксонами определяется не на основании филогении, а на основании математического анализа максимально большого количества признаков, имеющих одинаковое значение (вес).
Домены — относительно новый способ классификации. Трёхдоменная система изобретена в 1990 году, однако до сих пор не принята окончательно. Большинство биологов принимает эту систему доменов, однако значительная часть продолжает использовать пятицарственное деление. Одной из главных особенностей трёхдоменного метода является разделение археев(Archaea) и бактерий (Bacteria), которые ранее были объединены в царство бактерий. Существует также малая часть учёных, добавляющих археев в виде шестого царства, но не признающих домены.
Сегодня систематика принадлежит к числу бурно развивающихся биологических наук, включая всё новые и новые методы: методы математической статистики, компьютерный анализ данных, сравнительный анализ ДНК и РНК, анализ ультраструктуры клеток и многие другие.
[править]Эволюция систем классификации
Геккель (1894) Три царства |
Уиттекер (1969) Пять царств |
Вёзе (1977) Шесть царств |
Вёзе (1990) Три домена |
Кавалье-Смит (1998) Два домена и семь царств |
||
Животные |
Животные |
Животные |
Эукариоты |
Эукариоты |
Животные |
|
Растения |
Грибы |
Грибы |
Грибы |
|||
Растения |
Растения |
Растения |
||||
Простейшие |
Простейшие |
Хромисты |
||||
Протисты |
Протисты |
|||||
Монеры |
Археи |
- |
Археи |
Прокариоты |
Археи |
|
Бактерии |
Бактерии |
Бактерии |
||||
Вопрос №7
Медицинская протозоология |
|
Содержание |
1. Медицинская протозоология, предмет и задачи. 2. Общая характеристика типа Простейшие, принципы классификации. 3. Характеристика класса корненожек. 4. Строение, цикл развития дизентерийной амебы. 5. Характеристика класса жгутиковых. 6. Строение, цикл развития лямблии. 7. Характеристика класса инфузории. 8. Строение и цикл развития балантидия. 9. Диагностика и профилактика амебиаза, лямблиоза, балантидиаза. 10. Характеристика класса споровики. 11. Цикл развития возбудителя трехдневной малярии. 12. Диагностика и профилактика малярии. Список используемой литературы |
Введение |
Протозоология – это раздел зоологии, который занимается изучением всех видов простейших. К простейшим относятся животные, состоящие из одной или нескольких клеток. Каждая клетка – это самостоятельный орга-низм, даже если они и объединены в колонии. Простейшие ведут различный образ жизни. Огромное число их являются паразитами, то есть живут в теле других животных, нанося им вред. Простейшие, живущие в теле человека, могут являться возбудителями инфекционных заболеваний. Долгое время не было известно о настоящих причинах таких болезней. Только изобретение микроскопа позволило уви-деть микроорганизмы, живущие в организме человека и сделать вывод, что именно они являются причиной инфекционных заболеваний. Медицинская протозоология занимается изучением микроорганизмов, которые являются возбудителями инфекций. Предметом ее исследования являются животные типа Простейшие, вызывающие инфекционные заболе-вания, развитие микроорганизмов, вызывающих болезни. Такими формами микроорганизмов является класс споровиков, в который входят малярийные плазмодии, вызывающие малярию, корненожки, дизентерийные амебы. Ос-новной задачей медицинской протозоологии является диагностика, лечение и профилактика таких заболеваний. |
Заключение |
В настоящее время малярия как массовое заболевание ликвидирована, регистрируется единичные случаи болезни. При микроскопическом исследовании в крови больного малярией можно обнаружить плазмодии, расположенные внутри эритроцитов. Для трех-дневной малярии характерна форма плазмодия в виде кольца с камнем. Развитие плазмодиев в организме больного человека происходит глав-ным образом в результате их паразитирования внутри эритроцитов. Этому бесполому циклу предшествует образование тканевых форм (в селезенке, печени, костном мозгу) из спорозоитов, которые проникли в организм че-ловека со слюной зараженного комара анафелес при укусе им здорового че-ловека. Перед наступлением характерных лихорадочных приступов малярии не-редко наблюдается продромальный период, который длится 1-3 дня и вы-ражается в общем недомогании, слабости, ухудшении аппетита. При пер-вичном заражении развивается лихорадочная реакция с повышением тем-пературы. В течении 5-6 дней температура сохраняется на этом уровне, за-тем возникают периодические, следующие один за другим через опреде-ленные промежутки времени, приступы малярии. Приступ развивается обычно в утренние часы. Вначале проявляется сильный озноб, ощущение холода. Вслед за ознобом очень быстро нараста-ет температура. На высоте приступа, когда температура значительно повы-силась, больному становится жарко, его мучает сильная жажда. Кожа боль-ного сухая, горячая на ощупь, его мучает сильная жажда, нередко на коже губ имеются герпетические высыпания. При трехдневной малярии по прошествии 5-6 часов от начала приступа больной начинает сильно потеть. Затем температура критически падает до нормальных цифр, больной испытывает значительное облегчение Время от момента до окончания одного лихорадочного приступа до начала развития следующего носит название периода апирексии. При трехдневной лихорадке от начала одного приступа до начала друго-го проходит 48 часов. Как правило, больные малярией имеют плохой аппетит, во время при-ступа наблюдается давление в подложечной области, тошнота и рвота. При длительном течении болезни язык обычно обложен, живот увеличен в раз-мерах за счет увеличения печени и селезенки. Диагноз малярии основывается на данных эпидемиологии, клинической картине и течении болезни. Если в первые дни заболевания иногда трудно поставить диагноз, то с появлением характерных малярийных приступов он значительно облегчается. Может потребоваться проведение дифференци-ального диагноза с брюшным тифом, бруцеллезом, пневмонией. Профилактика направлена на раннее выявление и лечение больных, а также на борьбу с комарами – переносчиками малярии. Большое значение в борьбе с комарами имеет ликвидация мест их выплода, благоустройство хо-зяйственно-полезных водоемов. В период активности комаров проводится их истребление в помещениях, устанавливают марлевые или металлические сетки на окнах и дверях, для защиты от нападения комаров используют ре-пелленты. В местностях, где регистрируются заболевания малярией, при выезде в неблагополучные по малярии районы проводится предупредительное лече-ние. Необходимо также предпринимать меры по защите больных малярией от нападения комаров. Проводится также химпрофилактика малярии. Своевременное выявление больных малярией людей и их рациональное лечение являются важными мерами в комплексе средств борьбы с распро-странителем болезни. Систематическое лечение ведет к ликвидации носительства плазмодий. У каждого больного, подозрительного на заболевание малярией, особенно в весенне-летний период и в тех местностях, где встре-чается малярия, необходимо микроскопическое исследование крови. |
|
|
Вопрос №8
95 Простейшие. Классификация, характерные черты организации.
Наука о паразитических простейших - возбудителях болезней человека, носит название медицинская протозоология.
Морфологическая характеристика. Тело простейших имеет микроскопические размеры и представлено одной клеткой. Клетка эукариотическая. Органоиды простейших осуществляют функции целого организма.
Органоиды передвижения - псевдоподии, жгутики или реснички.
Органоиды питания - пищеварительные вакуоли. Непереваренные остатки пищи выбрасываются наружу.
Органоиды выделения и осморегуляции - сократительные вакуоли. У паразитических и морских форм этот тип вакуоли может отсутствовать.
Органоиды защиты - трихоцисты. Существу001ют у инфузорий.
Размножаются простейшие бесполым и половым путем. Бесполое размножение: бинарное и множественное (шизогония) деления. Половой процесс: копуляция и конъюгация.
Жизненный цикл. Стадии развития в цикле часто повторяются с определенной закономерностью: зигота, бесполое поколение, половое поколение и вновь зигота. Имеются активно питающиеся, подвижные стадии - трофозоиты. Простейшие способны к инцистированию. Среда обитания: свободноживущих - в пресной и морской воде, в почве, паразитических форм - в жидких средах и клетках организмов.
Классификация: 1. Класс Саркодовые - Sarcodina; 2. Класс Жгутиковые - Flagellata s. Mastigophora; 3. Класс Инфузории - Infusoria; 4. Класс Споровики - Sporozoa.
Паразитические виды встречаются в каждом классе.
ОБЩАЯ ХАРАКТЕРИСТИКА
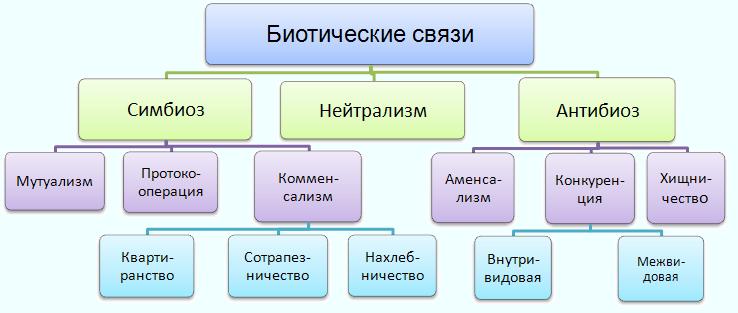
Первый тип, с которого мы начинаем знакомство с миром животных, — это тип простейших (Protozoa). Он состоит из многих классов, отрядов, семейств и включает примерно 20—25 тыс. видов.
Простейшие распространены на всей поверхности нашей планеты и живут в самых различных средах. В большом количестве мы найдем их в морях и океанах, как непосредственно в толще морской воды, так и на дне. Обильны простейшие в пресных водах. Некоторые виды живут в почве.
Многие простейшие в качестве среды обитания используют другие организмы, ведут паразитический образ жизни. Среди паразитических простейших имеются возбудители тяжелых заболеваний человека, домашних и промысловых животных.
По своему строению простейшие чрезвычайно разнообразны. Подавляющее большинство их обладает микроскопически малыми размерами, для изучения их приходится пользоваться микроскопом.
Каковы же общие признаки типа простейших? На основании каких особенностей строения и физиологии мы причисляем животных к этому типу? Основной и самой характерной чертой простейших является их одноклеточность. Простейшие являются организмами, тело которых по строению соответствует одной клетке.
Все другие животные (а также и растения) тоже состоят из клеток и их производных. Однако, в отличие от простейших, в состав тела их входит большое количество клеток, различных по строению и выполняющих в сложном организме разные функции. По этому признаку все остальные животные могут быть противопоставлены простейшим и отнесены к многоклеточным (Metazoa).
Сходные по строению и функции клетки их слагаются в комплексы, называемые тканями. Органы многоклеточных состоят из тканей. Различают, например, покровную (эпителиальную) ткань, мышечную ткань, нервную ткань и др.
Если по строению своему простейшие соответствуют клеткам многоклеточных организмов, то в функциональном отношении они несравнимы с ними. Клетка в теле многоклеточного всегда представляет собой только часть организма, ее отправления подчинены функциям многоклеточного организма как целого. Напротив, простейшее — это самостоятельный организм, которому свойственны все жизненные функции: обмен веществ, раздражимость, движение, размножение.
К окружающим условиям внешней среды простейшее приспосабливается как целый организм. Следовательно, можно сказать, что простейшее — это самостоятельный организм на клеточном уровне организации.
Размеры тела подавляющего большинства простейших микроскопические. Наиболее мелкие представители типа имеют 2—4 микрона в поперечнике (например, паразитические жгутиконосцы из рода лейшманий — Leichmania, разные виды семейства пироплазмид, паразитирующие в красных кровяных клетках млекопитающих).
Наиболее обычные размеры простейших — в пределах 50—150 мк. Но среди них имеются и гораздо более крупные организмы.
Инфузории Bursaria, Spirostomum достигают 1, 5 мм длины—их хорошо видно простым глазом, грегарины Porospora gigantea — длины до 1 см.
У некоторых корненожек фораминифер раковина достигает 5—6 см в диаметре (например, виды рода Psammonix, ископаемые нуммулиты и др.).
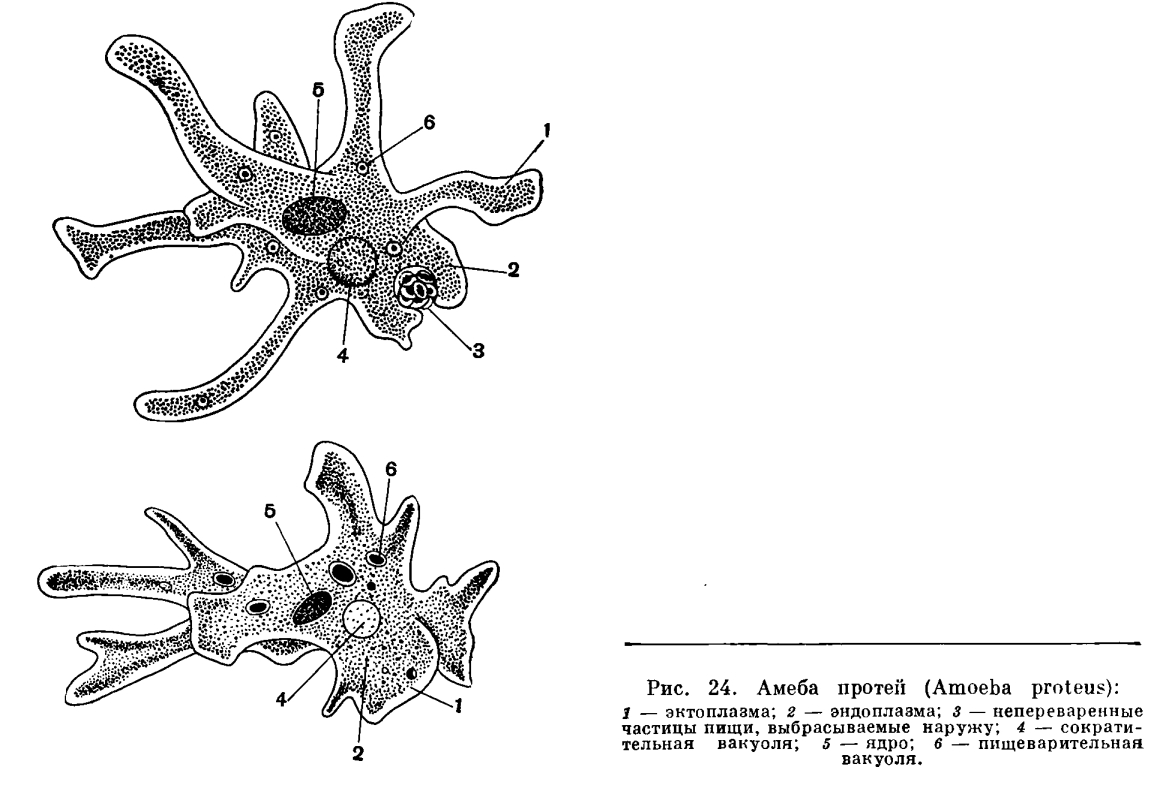
Низшие представители простейших (например, амебы) не обладают постоянной формой тела. Их полужидкая цитоплазма постоянно меняет свои очертания благодаря образованию разнообразных выростов — ложных ножек (рис. 24), служащих для движения и захвата пищи.
Большинство же простейших обладает относительно постоянной формой тела, которая обусловлена наличием опорных структур. Среди них наиболее обычной является плотная эластичная мембрана (оболочка), образуемая периферическим слоем цитоплазмы (эктоплазмой) и носящая название пелликулы.
В одних случаях пелликула относительно тонка и не препятствует некоторому изменению формы тела простейшего, как это имеет место, например, у способных сокращаться инфузорий. У других простейших она образует прочный и не меняющий своей формы наружный панцирь.
У многих жгутиконосцев, окрашенных в зеленый цвет благодаря наличию хлорофилла, имеется наружная оболочка из клетчатки — признак, характерный для растительных клеток.
Что касается общего плана строения и элементов симметрии, то простейшие обнаруживают большое разнообразие. Такие животные, как амебы, не обладающие постоянной формой тела, не имеют постоянных элементов симметрии.
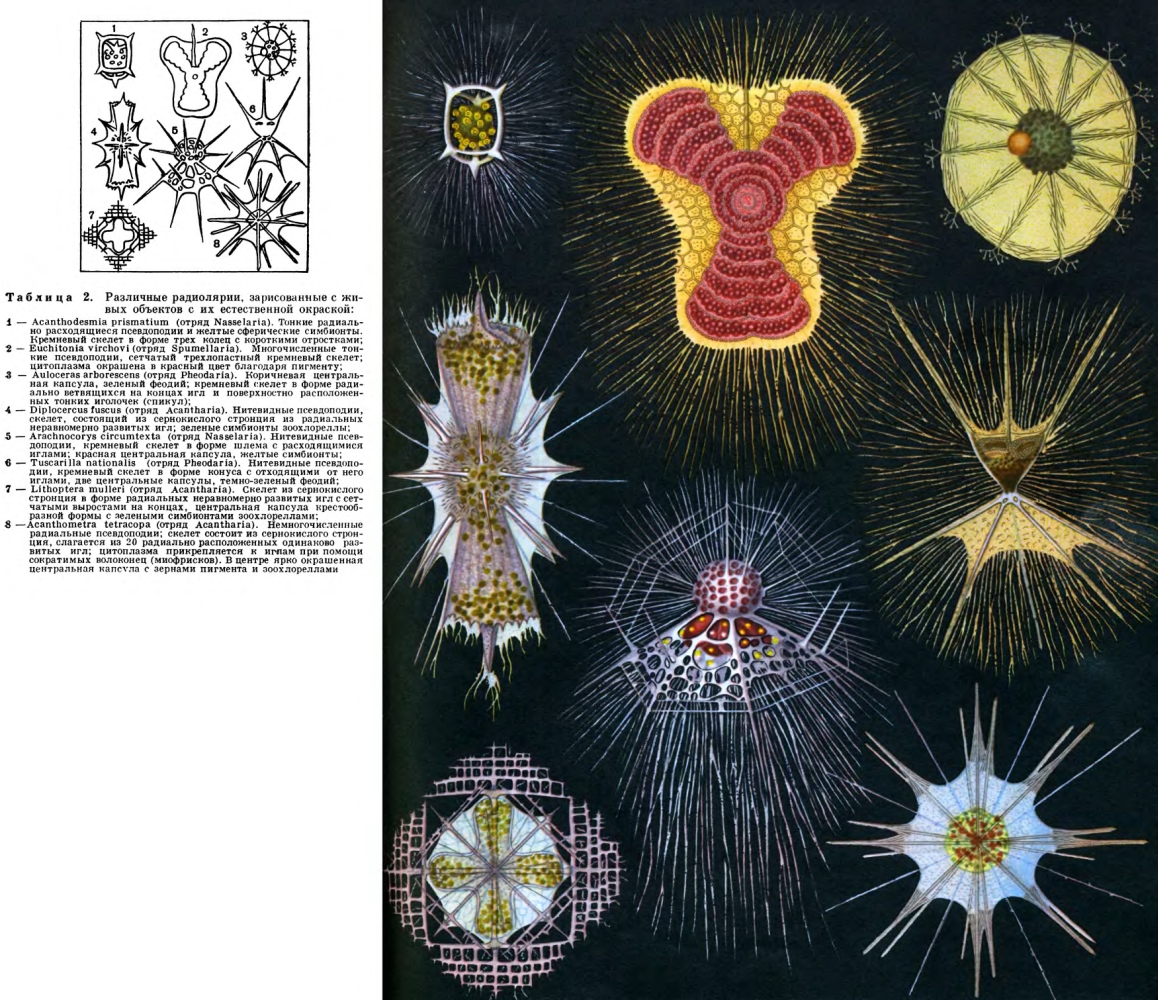
Широко распространены среди Protozoa разные формы радиальной симметрии, свойственной главным образом планктонным формам (многие радиолярии, солнечники). При этом имеется один центр симметрии, от которого отходит различное число пересекающихся в центре осей симметрии, определяющих расположение частей тела простейшего.
,
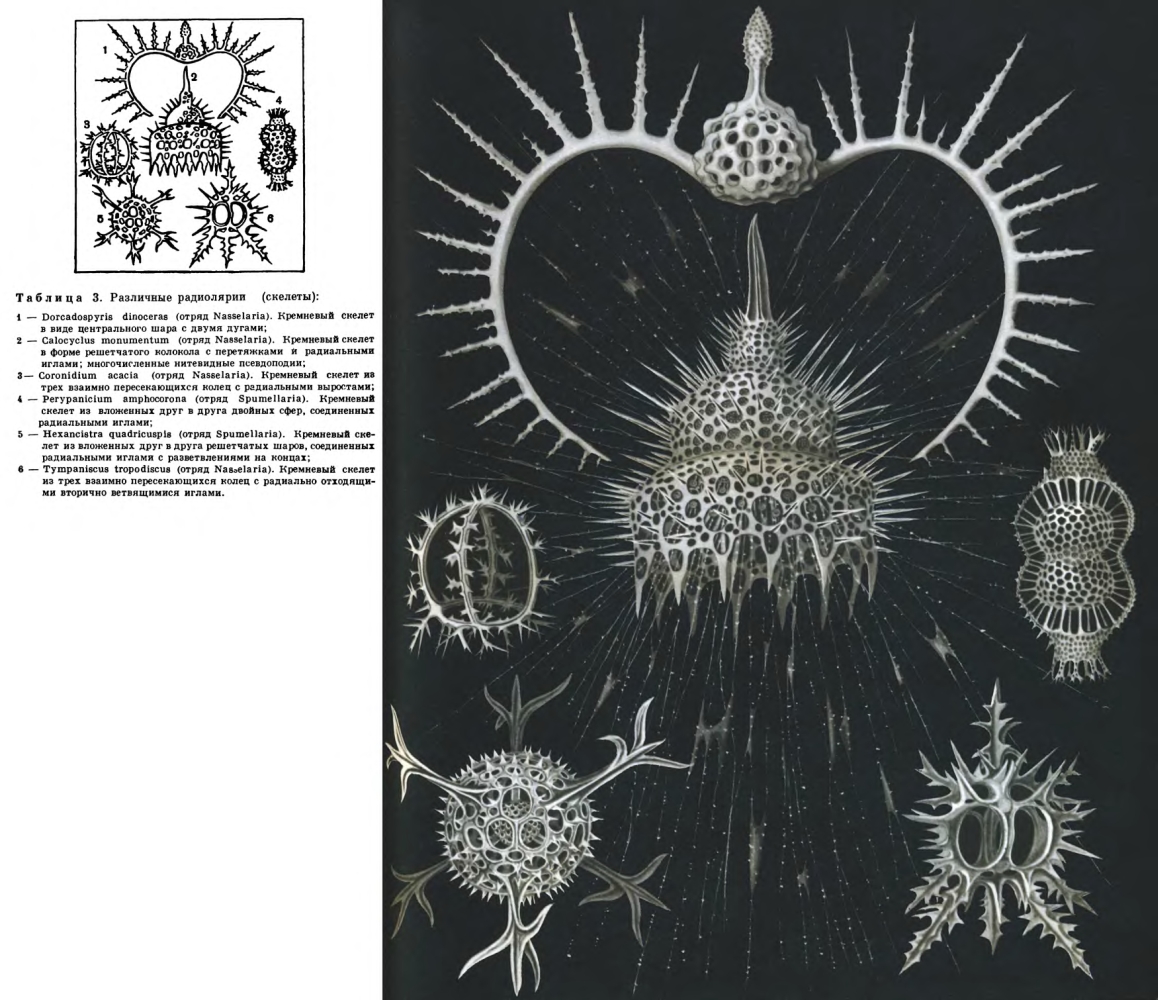
У многих радиально построенных форм можно выделить одну главную ось, определяющую передний и задний концы тела, вокруг которой радиально располагаются части тела простейшего (некоторые радиолярии, табл. 2, 3, инфузории Didinium).

,

,
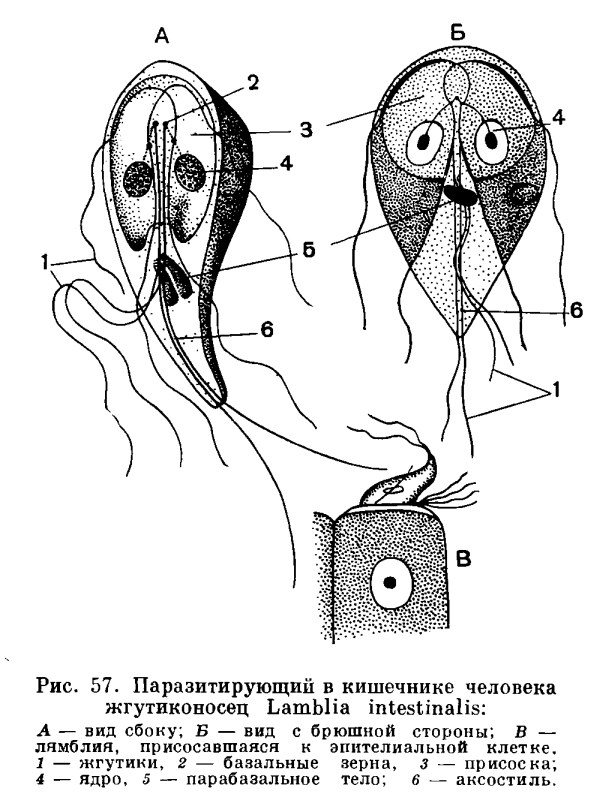
Относительно редко встречается у простейших двубоковая (билатеральная) симметрия, при которой можно провести одну-единственную плоскость симметрии, делящую тело животного на две равные зеркальные половины (раковины некоторых фораминифер, рис. 32, 33, радиолярии, табл. 2 и 3, некоторые виды жгутиконосцев, например лямблия, рис. 57). Большинство простейших из разных классов являются асимметричными.
У сложно организованных простейших из класса инфузорий и у некоторых жгутиконосцев, кроме пелликулы, имеются еще и другие опорные структуры, поддерживающие и определяющие форму тела. К ним относятся тончайшие волоконца (фибриллы), проходящие в различных направлениях. Примером могут служить опорные волоконца одной из инфузорий.
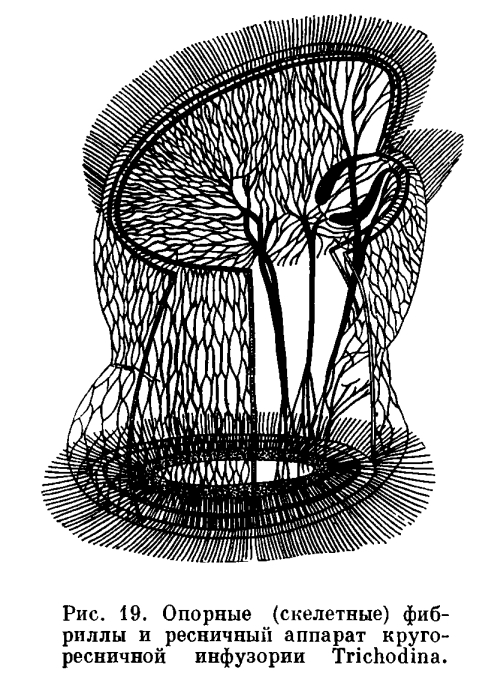
На рисунке 19 видно, какой большой сложности может достигать эта система, образующая прочный и эластичный каркас, поддерживающий полужидкую цитоплазму простейшего.
К числу опорных и вместе с тем защитных образований у простейших относятся различные формы минерального скелета, свойственного преимущественно многим представителям класса саркодовых. Эти скелетные образования чаще всего имеют форму раковинок, иногда очень сложно устроенных (в отряде фораминифер). В других случаях основу скелета составляют отдельные иглы (спикулы), обычно соединяющиеся между собой, По химическому составу минеральный скелет простейших различен. Наиболее обычными компонентами его являются углекислый кальций (СаС03) или окись кремния (Si02). Более подробно строение скелета будет рассмотрено при знакомстве с отдельными классами простейших.
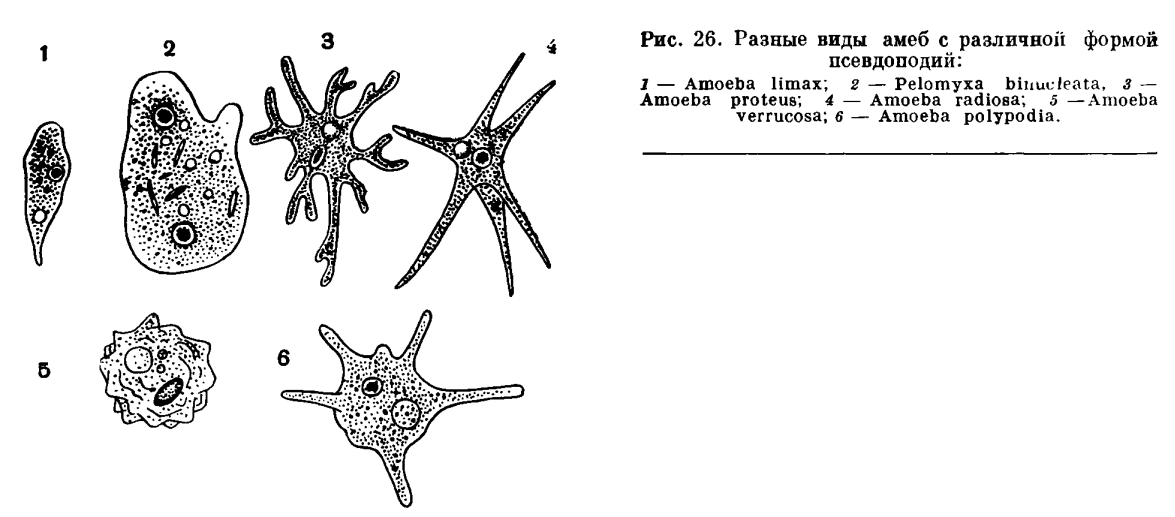
Большинству простейших, за исключением некоторых паразитических форм, свойственна способность к движению — перемещению в пространстве. Формы движения простейших разнообразны. Наиболее простой и, вероятно, исходной формой является амебоидное движение. Оно выражается в образовании ложных ножек (псевдоподий) — выростов цитоплазмы разной формы. Все содержимое клетки как бы медленно перетекает в направлении образующейся псевдоподии, и таким путем осуществляется перемещение простейшего в пространстве. Эта форма движения преимущественно свойственна представителям класса саркодовых. Разным видам свойственна различная форма псевдоподий (рис. 26).
Более сложной формой является движение, осуществляемое при помощи жгутиков и ресничек. Жгутиковая форма движения характерна для класса жгутиконосцев.
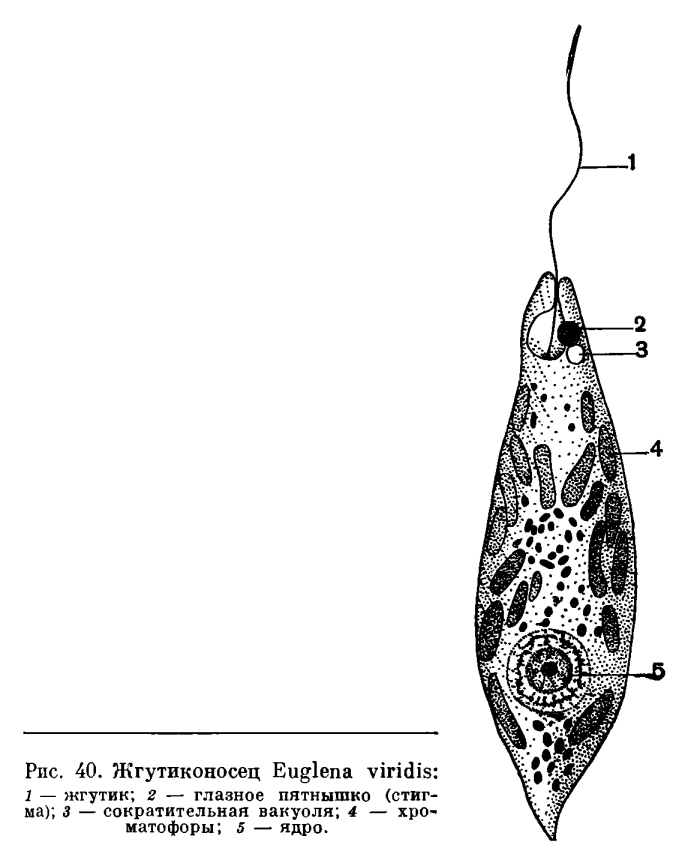
,
Жгутики представляют собой тончайшие выросты тела. Количество их у разных видов различно — от одного до многих десятков и даже сотен (рис. 40, 63). Каждый жгутик берет начало от небольшого базального зернышка, называемого блефаропластом и расположенного в цитоплазме. Таким образом, непосредственно граничащая с базальным зерном часть жгутика проходит внутри цитоплазмы (она носит название корневой нити), а затем проходит через пелликулу наружу. Механизм жгутикового движения у разных видов различен. В большинстве случаев он сводится к вращательному движению. Жгутик описывает фигуру конуса, вершиной обращенного к месту его прикрепления. Наибольший механический эффект достигается, когда угол, образуемый вершиной конуса, составляет 40—46°. Быстрота движения различна, она колеблется у разных видов между 10 и 40 оборотами в секунду. Простейшее как бы «ввинчивается» в окружающую его жидкую среду.
Нередко вращательное движение жгутика сочетается с его волнообразным движением. Обычно при поступательном движении само тело простейшего вращается вокруг продольной оси.
Изложенная схема справедлива для большинства одножгутиковых форм. У многожгутиковых движение жгутиков может носить иной характер, в частности жгутики могут находиться в одной плоскости, не образуя конуса вращения.
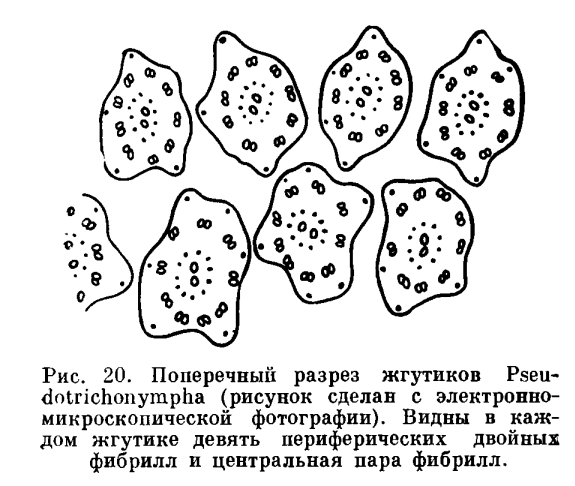
Электронномикроскопические исследования последних лет показали, что внутренняя ультрамикроскопическая структура жгутиков весьма сложна. Снаружи жгутик окружен тонкой мембраной, которая является непосредственным продолжением самого поверхностного слоя эктоплазмы — пелликулы. Внутренняя полость жгутика заполнена цитоплазматическим содержимым. По продольной оси жгутика проходит одиннадцать тончайших нитей (фибрилл), которые нередко являются двойными (рис. 20). Эти фибриллы располагаются всегда закономерно. Девять из них (простых или двойных) лежат по периферии, образуя в совокупности как бы цилиндр. Две фибриллы занимают центральное положение. Чтобы составить себе представление о размерах всех этих образований, достаточно сказать, что диаметр периферических фибрилл составляет около 350А (ангстрем). Ангстрем — единица длины, равная 0, 0001 мк, а микрон равняется 0, 001 мм. Вот какие ничтожные по своим размерам структуры стали доступными для изучения благодаря внедрению в микроскопическую технику электронного микроскопа.
Функциональное значение фибрилл жгутиков не может считаться окончательно выясненным. По-видимому, часть их (вероятно, периферические) играет активную роль в двигательной функции жгутика и содержит особые белковые молекулы, способные сокращаться, другие же являются опорными эластическими структурами, имеющими поддерживающее значение.
Реснички служат органоидами движения инфузорий. Обычно число их у каждой особи очень велико и измеряется несколькими сотнями, тысячами и даже десятками тысяч. Механизм движения ресничек несколько иной, чем жгутиков. Каждая ресничка совершает гребные движения. Она быстро и с силой сгибается в одну сторону, а затем медленно выпрямляется.
Совместное действие большого числа ресничек, биение которых координировано, вызывает быстрое поступательное движение простейшего.
Каждая ресничка инфузории, как показали новейшие исследования, является сложным образованием, по своему строению соответствующим жгутику. У основания каждой реснички всегда располагается так называемое базальное зерно (иначе, кинетозома) — важная часть ресничного аппарата.
У многих инфузорий отдельные реснички соединяются друг с другом, образуя структуры более сложного строения (мембранеллы, цирры и др.) и более эффективного механического действия.
Некоторым высокоорганизованным простейшим (инфузориям, радиоляриям) свойственна еще одна форма движения — сокращение. Тело таких простейших способно быстро менять свою форму, а затем вновь возвращаться к исходному состоянию.
Способность к быстрому сокращению обусловлена наличием в теле простейшего особых волоконец — мионем — образований, аналогичных мышцам многоклеточных животных.
У некоторых простейших существуют еще и другие формы движения.
По способам и характеру питания, по типу обмена веществ простейшие обнаруживают большое разнообразие.
В классе жгутиконосцев имеются организмы, способные подобно зеленым растениям при участии зеленого пигмента хлорофилла усваивать неорганические вещества — углекислый газ и воду, превращая их в органические соединения (аутотрофный тип обмена). Этот процесс фотосинтеза протекает с поглощением энергии. Источником последней является лучистая энергия — солнечный луч.
Таким образом, эти простейшие организмы правильнее всего рассматривать как одноклеточные водоросли. Но наряду сними в пределах того же класса жгутиконосцев имеются бесцветные (лишенные хлорофилла) организмы, неспособные к фотосинтезу и обладающие гетеротрофным (животным) типом обмена веществ, т. е. питающиеся за счет готовых органических веществ. Способы животного питания простейших, так же как и характер пищи их, очень разнообразны. Наиболее просто устроенные простейшие не обладают специальными органоидами захвата пищи. У амеб, например, псевдоподии служат не только для движения, но вместе с тем и для захвата оформленных частиц пищи. У инфузорий для захвата пищи служит ротовое отверстие. С последним обычно связаны разнообразные структуры — околоротовые мерцательные перепонки (мембранеллы), способствующие направлению пищевых частиц к ротовому отверстию и далее в особую трубку, ведущую в эндоплазму — клеточную глотку.
Пища простейших очень разнообразна. Одни питаются мельчайшими организмами, например бактериями, другие — одноклеточными водорослями, некоторые являются хищниками, пожирающими других простейших, и т. п. Непереваренные остатки пищи выбрасываются наружу — у саркодовых на любом участке тела, у инфузорий через особое отверстие в пелликуле.
У пресноводных и у части морских простейших имеется особый органоид, связанный с регуляцией осмотического давления в клетке простейшего и с выделением. Это сократительная вакуоля. Она представляет собой периодически появляющийся в цитоплазме пузырек, наполняющийся жидкостью и опорожняющийся наружу. Сократительная вакуоля регулирует количество воды в цитоплазме, поступающей из окружающей среды благодаря разности осмотических давлений. У паразитических простейших и у многих морских форм, живущих в среде с повышенным осмотическим давлением, сократительные вакуоли отсутствуют.
Особых органоидов дыхания у простейших нет, они поглощают кислород и выделяют углекислоту всей поверхностью тела.
Как и все живые существа, простейшие обладают раздражимостью, т. е. способностью отвечать той или иной реакцией на факторы, действующие извне. Простейшие реагируют на механические, химические, термические, световые, электрические и иные раздражения. Реакции простейших на внешние раздражения часто выражаются в изменении направления движения и носят название таксис. Таксисы могут быть положительными, если движение осуществляется в направлении раздражителя, и отрицательными, если оно осуществляется в противоположную сторону.
Реакции многоклеточных животных на раздражения осуществляются под воздействием нервной системы. Многие исследователи пытались обнаружить и у простейших (т. е. в пределах клетки) аналоги нервной системы. Американские ученые, например, описывали у многих инфузорий наличие особого нервного центра (так называемого моториума), представляющего собой особый уплотненный участок цитоплазмы. От этого центра к различным участкам тела инфузории отходит система тонких волоконец, которые рассматривались как проводники нервных импульсов. Другие исследователи, применяя особые методы серебрения препаратов (обработка азотнокислым серебром с последующим восстановлением металлического серебра), обнаружили в эктоплазме инфузорий сеть тончайших волоконец. Эти структуры (рис. 21) также рассматривались как нервные элементы, по которым распространяется волна возбуждения. В настоящее время, однако, большинство ученых, изучающих тонкие фибриллярные структуры, придерживаются иного мнения об их функциональной роли в клетке простейшего. Экспериментальных доказательств нервной роли фибриллярных структур не получено. Напротив, имеются опытные данные, которые дают возможность предполагать, что у простейших волна возбуждения распространяется непосредственно по наружному слою цитоплазмы — эктоплазме. Что же касается различного рода фибриллярных структур, которые еще недавно рассматривались как «нервная система» простейших, то они имеют скорее всего опорное (скелетное) значение и способствуют сохранению формы тела простейшего.
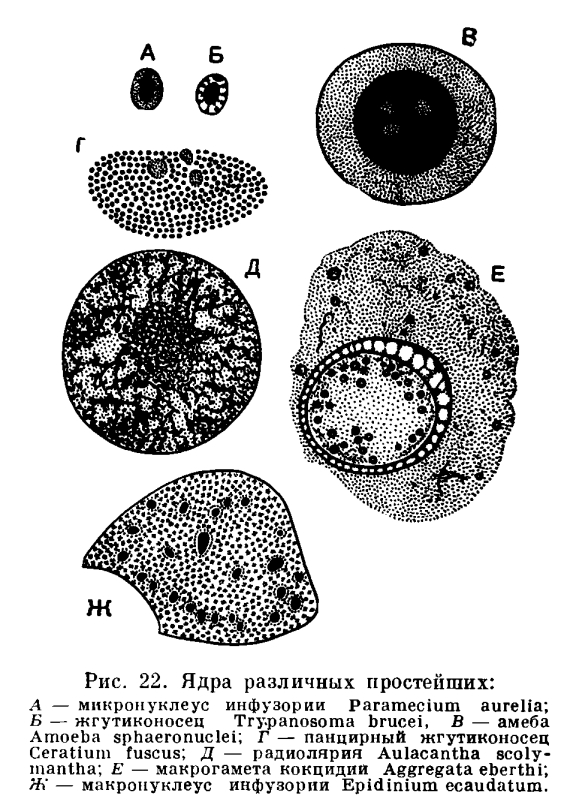
Как н всякая клетка, простейшие имеют ядро. Выше, при рассмотрении строения клетки, мы уже ознакомились с основными структурными компонентами ядра. В ядрах простейших, так же как и в ядрах многоклеточных, имеется оболочка, ядерный сок (кариолимфа), хроматин (хромосомы) и ядрышки. Однако по размерам и строению ядра разные простейшие весьма разнообразны (рис. 22). Эти различия обусловлены соотношением структурных компонентов ядра: количеством ядерного сока, количеством и размерами ядрышек (нуклеол), степенью сохранения строения хромосом в интерфазном ядре и т. п.
У большинства простейших имеется одно ядро. Однако встречаются и многоядерные виды простейших.
У некоторых простейших, а именно у инфузорий и немногих корненожек — фораминифер, наблюдается интересное явление дуализма (двойственности) ядерного аппарата. Оно сводится к тому, что в теле простейшего имеются два ядра двух категорий, различающиеся как по своему строению, так и по физиологической роли в клетке. У инфузорий, например, имеется два типа ядер: большое, богатое хроматином ядро — макронуклеус и маленькое ядро — микронуклеус. Первое связано с выполнением вегетативных функций в клетке, второе—с половым процессом.
Простейшим, как и всем организмам, свойственно размножение. Существуют две основные формы размножения простейших: бесполое и половое. В основе того и другого лежит процесс деления клетки.
При бесполом размножении число особей возрастает в результате деления. Например, амеба при бесполом размножении делится на две амебы путем перетяжки тела. Процесс этот начинается с ядра, а затем захватывает цитоплазму. Иногда бесполое размножение приобретает характер множественного деления. При этом ядро предварительно делится несколько раз и простейшее становится многоядерным. Вслед за этим цитоплазма распадается на число отдельностей, соответствующих количеству ядер. В результате организм простейшего сразу дает начало значительному количеству мелких особей. Так происходит, например, бесполое размножение малярийного плазмодия — возбудителя малярии человека.
Половое размножение простейших характеризуется тем, что собственно размножению (увеличению числа особей) предшествует половой процесс, характерным признаком которого является слияние двух половых клеток (гамет) или двух половых ядер, ведущее к образованию одной клетки — зиготы, дающей начало новому поколению. Формы полового процесса и полового размножения у простейших в высшей степени разнообразны. Основные формы его будут рассмотрены при изучении отдельных классов.
Многие виды простейших имеют не одну, а несколько форм размножения, которые могут закономерно чередоваться друг с другом. В результате получается сложный цикл развития, отдельные этапы которого могут протекать в разных условиях среды. Особенной сложностью отличаются циклы развития у паразитических простейших в классе споровиков.
Простейшие обитают в самых различных условиях среды. Большинство их — водные организмы, широко распространенные как в пресных, так и в морских водоемах. Многие виды их живут в придонных слоях и входят в состав бентоса. Большой интерес представляет приспособление простейших к жизни в толще песка, в толще воды (планктон).
Небольшое число видов Protozoa приспособилось к жизни в почве. Их средой обитания являются тончайшие пленки воды, окружающие почвенные частицы и заполняющие капиллярные просветы в почве. Интересно отметить, что даже в песках пустыни Каракум живут простейшие. Дело в том, что под самым верхним слоем песка здесь расположен влажный слон, пропитанный водой, приближающейся по своему составу к морской воде. В этом влажном слое и были обнаружены живые простейшие из отряда фораминифер, являющиеся, по-видимому, остатками морской фауны, населявшей моря, ранее находившиеся на месте современной пустыни. Эта своеобразная реликтовая фауна в песках Каракумов впервые была обнаружена проф. Л. Л. Бродским при изучении воды, взятой из колодцев пустыни.
Очень многие простейшие, относящиеся к различным классам этого типа, ведут паразитический образ жизни. Их среда обитания и источники пищи — другие живые организмы. Сыше 3, 5 тыс. видов простейших — паразиты. Хозяевами их являются самые различные животные и растения. Многие паразитические простейшие живут в организме человека, домашних и промысловых животных, нанося им большой вред. Один из классов типа простейших — споровики — целиком состоит из паразитов.
Практическое значение простейших для человека велико. В особенности это относится к паразитам. До настоящего времени в тропических зонах земного шара широко распространена малярия — тяжелое заболевание, поражающее ежегодно десятки миллионов человек в Индии и других тропических частях Азии, в Африке и других странах. Возбудитель этого заболевания относится к классу споровиков типа простейших. Тяжелым заболеванием человека в Центральной Африке является сонная болезнь, вызываемая паразитом из класса жгутиконосцев. Большой ущерб наносят животноводству заболевания до машних животных, вызываемые простейшими. Сюда относятся различные пироплазмозы, кокцидиозы, трипанозомозы и многие другие. Важнейшие из них в практическом отношении мы рассмотрим ниже.
Имеется ряд простейших паразитов из отряда кощидий, поражающих домашнюю птицу, в особенности кур. Борьба с этими многочисленными и опасными протозойными заболеваниями требует детального изучения биологии возбудителей, их циклов развития.
Некоторый практический интерес представляют и свободноживущие простейшие. Разные виды их приурочены к определенному комплексу внешних условий, в частности к различному химическому составу воды.
Определенные виды простейших живут при разной степени загрязненности пресных вод органическими веществами. Поэтому по видовому составу простейших можно судить о свойствах воды водоема. Эти особенности простейших используют для санитарно-гигиенических целей при так называемом биологическом анализе воды.
В общем круговороте веществ в природе простейшие играют заметную роль. В водоемах многие из них являются энергичными пожирателями бактерий и других микроорганизмов. Вместе с тем сами они служат пищей для более крупных животных организмов. В частности, выклевывающиеся из икринок мальки многих видов рыб на самых начальных этапах своей жизни питаются преимущественно простейшими.
Тип простейших в геологическом отношении является весьма древним. В ископаемом состоянии хорошо сохранились те виды простейших, которые обладали минеральным скелетом (фораминиферы, радиолярии). Ископаемые остатки их известны начиная с самых древних нижнекембрийских отложений.
Морские простейшие — корненожки и радиолярии — играли и играют весьма существенную роль в образовании морских осадочных пород. В течение многих миллионов и десятков миллионов лет микроскопически мелкие минеральные скелеты простейших после отмирания животных опускались на дно, образуя здесь мощные морские отложения. При изменении рельефа земной коры, при горнообразовательных процессах в прошлые геологические эпохи, морское дно становилось сушей. Морские осадки превращались в осадочные горные породы. Многие из них, как, например, некоторые известняки, меловые отложения и др., в значительной своей части состоят из остатков скелетов морских простейших. В силу этого изучение палеонтологических остатков простейших играет большую роль в определении возраста разных слоев земной коры и, следовательно, имеет существенное значение при геологической разведке, в частности при разведке полезных ископаемых.
ИСТОРИЯ ИЗУЧЕНИЯ ПРОСТЕЙШИХ
Изучение простейших началось значительно позже, чем изучение большинства других групп животного мира. Оно стало возможным лишь после изобретения микроскопа, что произошло в начале XVII в.
В 1675 г. голландец Антон Левенгук, рассматривая под микроскопом каплю воды, впервые открыл в ней множество микроскопических, ранее неведомых организмов, среди которых были и простейшие. Наблюдения Левенгука возбудили большой интерес к этому новому миру живых существ. В конце XVII и первой половине XVIII в. появляется большое число сочинений, посвященных изучению микроскопических организмов. Однако современного представления о простейших как одноклеточных организмах тогда не существовало, ибо само понятие о клетке было сформулировано лишь в конце первой половины XIX в. К этому вновь открытому миру микроскопических живых существ, которых чаще всего называли «мелкими наливочными животными» (Animalcula infusoria), относили самые различные организмы (простейших, круглых и ресничных червей, коловраток, одноклеточные водоросли ит. п.) по признаку их микроскопических размеров. Термин «инфузории», который в настоящее время обозначает один из классов простейших, в XVII—XVIII вв. имел совсем другое значение. Микроскопические организмы обильно развиваются в разных растительных настойках — infusum. Отсюда и произошло название, которое сначала не было связано с систематическим положением организмов, а означало «наливочные» или «настоечные» животные, т. е. развивающиеся в настойках.
Представления о строении и жизни микроскопических существ в XVII—XVIII вв., несмотря на большое количество посвященных им сочинений, были крайне неопределенными и хаотичными. Это дало знаменитому систематику Карлу Линнею основание для объединения в своей «Системе природы» (издание 1759 г.) всех известных ему простейших в один род, который он назвал весьма выразительно — Chaos infusorium.
Большое значение для познания микроскопических существ имело сочинение О. Ф. Мюллера «Animalcula infusoria» (1770), в котором описано 377 видов микроскопических организмов, главным образом простейших. Многие предложенные им родовые и видовые названия сохранились и в современной системе простейших. Мюллера нередко называют «Линнеем протистов», подчеркивая этим то большое значение, которое имели его работы для изучения мира микроскопических организмов.
Воззрения ученых на простейших в XVIII и начале XIX в. носили еще крайне противоречивый и подчас даже диаметрально противоположный характер. Так, например, Эренберг в своем известном сочинении «Наливочные животные как совершенные организмы» (1838) описывает простейших как сложно организованных существ, обладающих различными системами органов и отличающихся от других животных лишь своими размерами.
В противоположность Эренбергу другой крупный ученый этого периода, Дю-жардэн, в ряде сочинений утверждает, что простейшие не обладают никакой внутренней организацией и построены из бесструктурного полужидкого живого вещества — саркоды.
Название типа Protozoa впервые было введено в науку Гольдфусом в 1820 г. Однако наряду с простейшими в современном понимании к Protozoa он относил коловраток, мшанок, гидроидных полипов.
Понадобились еще многие годы работы, прежде чем удалось выяснить истинную природу простейших. Это стало возможным лишь после того, как в конце 30-х годов XIX в. трудами Шлейдена, Шванна и ряда других ученых было разработано учение о клетке.
Впервые в 1845 г. Зибольдом и Кёлликером было сформулировано представление о простейших как об одноклеточных организмах. Таким образом, тип Protozoa был четко отграничен от других типов микроскопических животных.
Для того чтобы определить границы типа и природу Protozoa, понадобилось 200 лет (со времен Левенгука) напряженных исследований.
Во второй половине XIX в. в изучении простейших особенно большую роль сыграли исследования немецкого биолога Бючли и его многочисленных учеников. Они изучили с позиций клеточной теории основные черты строения простейших и положили начало исследованию форм их размножения. Особенно большую роль для изучения половых процессов в размножении инфузорий сыграли работы Мопа.
В конце XIX и в первой четверти XX в. внимание исследователей привлекает изучение паразитических простейших и их циклов развития, что связано с большим практическим значением Protozoa как возбудителей опасных заболеваний человека и животных. В самом начале XX в. Шаудином (Германия) был впервые описан жизненный цикл кокцидий. Трудами Грасси (Италия), Данилевского (Россия) и других раскрыт жизненный цикл кровяных споровиков, что имело значение для борьбы с малярией.
В XX в. изучение простейших развивается очень быстро, что, в частности, связано с разработкой новых методов исследования их строения и физиологии: изучается размножение простейших из разных групп, физиологическая роль половых процессов (Калкинс, Вудруф, Дженнингс — США; Гертвиг — Германия; Метальников — Россия); исследуется изменчивость и наследственность; разрабатываются проблемы экологии и т. п. Изучение Protozoa все более и более тесно переплетается с проблемами исследования клетки (цитологии) и общей биологии.
За последние годы широкое применение в изучении простейших нашли методы электронной микроскопии, цитохимии, ультрафиолетовой микроскопии и др., о которых уже говорилось выше.
Русские и советские ученые внесли значительный вклад в изучение простейших. В конце XIX и начале XX в. профессор Петербургского университета Шевяков опубликовал ряд крупных исследований по инфузориям и радиоляриям. Особенно большой вклад в изучение систематики, строения, размножения и жизненных циклов простейших в течение второй четверти XX в. был внесен В. А. Догелем и его многочисленными учениками — протозоологами.
В области медицинской протозоологии (протозоология — область зоологии, изучающая простейших) большое значение имеют труды Данилевского, Марциновского, Эпштейна, Филипченко; в области ветеринарной протозоологии— Якимова, Маркова и многих других.
В настоящее время существует несколько международных научных журналов, где печатаются работы, посвященные изучению простейших. В ряде стран, втом числе и в Советском Союзе, изданы большие руководства, освещающие разные стороны протозоологии.
В 1961 г. в Праге состоялся первый международный конгресс протозоологов, на котором из всех стран мира собрались ученые, изучающие простейших. Второй международный конгресс протозоологов проходил в Лондоне в 1965 г.
Тип простейших (Protozoa) состоит из 5 классов: Саркодовые (Sarcodina), Жгутиконосцы (Mastigophora), Споровики (Sporozoa),Книдоспоридии (Cnidosporidia) и Инфузории (Infusoria).
Вопрс 9
Представители этого класса — самые примитивные простейшие. Форма их тела непостоянна. Передвигаются они с помощью ложноножек. Обитают в пресных водах, в почве, морях. В биогеоценозах выполняют функции консументов и редуцентов. Некоторые саркодовые адаптировались к комменсальному и паразитическому образу жизни. Медицинское значение имеют представители отряда амеб Amoebina. Паразитические амебы обитают у человека в основном в пищеварительной системе. Некоторые саркодовые, ведущие свободный образ жизни и обитающие в почве и загрязненной воде, при попадании в организм человека могут вызывать тяжелые заболевания, нередко заканчивающиеся смертью.
АМЕБА ДИЗЕНТЕРИЙНАЯ - Entamoeba histolytica - возбудитель кишечного (амебной дизентерии) и внекишечного амебиаза - антропоноза.
Географическое распространение - повсеместно, особенно часто в странах с тропическим и субтропическим климатом.
Локализация - слепая, восходящая, поперечно - ободочная кишка, а также печень, легкие, кожа и др.
Морфологическая характеристика. Существует в 4-х вегетативных формах - трофозоитах и цистной форме. 1. Мелкая вегетативная - просветная форма (f. minuta) (15-20 мкм) - непатогенна. У этой формы эктоплазма слабо выражена, движение медленное.
2. Тканевая форма (20 - 25 мкм) - патогенна. У амебы эктоплазма выражена, глыбки хроматина расположены радиально на периферии ядра, кариосома - строго в центре ядра, движение активное и сравнительно быстрое.
3. Крупная вегетативная (f. magna) (30 - 40 мкм до 60 - 80 мкм) - эритрофаг. Движение амебы активное, как у тканевой формы. При особых условиях (изменение бактериальной флоры кишечника, ослабление иммунитета) образует тканевую форму. При излечении заболевания эритрофаг переходит в просветную, а затем в предцистную форму. 4. Предцистная форма (12-20 мкм), ее цитоплазма не дифференцирована на экто- и эдоплазму, движение медленное. 5. Цистная форма (9 - 14 мкм) округлая с 4-мя ядрами. Незрелые цисты содержат овальные хроматоидные тельца. В зрелых цистах их нет.
Цикл развития.
Инвазионная форма - циста попадает к человеку через рот. Заражение цистами и просветными формами может сопровождаться бессимптомным носительством, чаще в средних широтах.
Патогенное действие. Патогенные формы вызывают изъязвления кишечника. Осложнения амебиаза: кишечное кровотечение и развитие абсцессов в печени. Редко может развиться абсцесс в легких, мозге, коже и других органах.
Источник заражения - больной человек и носитель.
Диагностика. Обнаружение тканевой и крупной вегетативных форм в мазке свежевзятых фекалий. Наличие просветных форм и цист недостаточно для диагностики амебиаза.
Эти формы как правило указывают на носительство. Возможна иммунологическая диагностика.
Профилактика: а) общественная - выявление и лечение больных и носителей; б) личная - соблюдение правил личной гигиены (мытье рук, овощей, фруктов, кипячение воды).
Дизентерийная амёба (лат. Entamoeba histolytica) — вид паразитических протозоа класса саркодовые. Вызывает тяжёлое заболевание —амебиаз (амёбную дизентерию, амёбный колит). Вид впервые описан в 1875 году русским ученым Ф. А. Лешем[1].
Размером дизентерийная амёба мельче обыкновенной амёбы (Amoeba proteus), подвижна. Эктоплазма чётко отграничена от эндоплазмы,псевдоподии короткие и широкие.
Содержание [убрать] 1 Жизненный цикл 1.1 Заражение 1.2 Просветная форма 1.3 Тканевая форма 1.4 Большая вегетативная форма 1.5 Циста 2 Примечания |
[править]Жизненный цикл
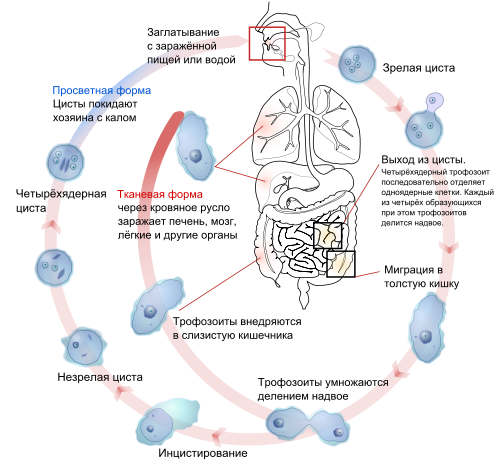
Может существовать в трёх формах: тканевой, просветной и цисты. Тканевая форма обнаруживается только у больных амёбиазом, другие — и у носителей.
[править]Заражение
Заражение наступает при попадании цист в верхний отдел толстого кишечника (слепая и восходящая ободочная кишка). Здесь цисты превращаются в просветные формы и внедряются в ткань кишки (тканевая форма), что сопровождается воспалением и формированием язв.
[править]Просветная форма
Просветная форма дизентерийной амёбы (лат. forma minuta) имеет размер около 20 мкм. Находится в верхнем отделе толстого кишечника. Движется с помощью псевдоподий. Ядросферическое, 3-5 мкм в поперечнике, хроматин расположен под ядерной оболочкой в виде небольших глыбок; в центре ядра небольшая кариосома.
[править]Тканевая форма
При внедрении просветной формы амёбы в ткани образуется тканевая форма (лат. forma magna) размером 20-60 мкм. В отличие от просветной формы не содержит в цитоплазме никаких включений. В этой стадии амёба размножается в стенке толстой кишки, образуя язвы. Язвенное поражение толстого кишечника сопровождается выделением слизи, гноя и крови.
[править]Большая вегетативная форма
Просветные и тканевые формы амёбы, попавшие в просвет кишки из язв, увеличиваются в размере до 30 мкм и больше и приобретают способность фагоцитировать эритроциты. Эта форма называется большой вегетативной, или эритрофагом.
Иногда амёбы из кишечника по кровеносным сосудам проникают в другие органы (прежде всего печень), формируя там вторичные очаги — абсцессы (внекишечный амёбиаз).
При затихании острой фазы болезни большая вегетативная форма уменьшается в размерах, переходит в просветную форму, которая инцистируется в кишечнике. Выброшенная при дефекации во внешнюю среду, она погибает в течение 15-20 минут.
[править]Циста
![]()
Циста
Цисты образуются при сгущении фекалий в толстой кишке. Просветная форма окружается оболочкой и превращается в шаровидную цисту (размер около 12 мкм) с 4 ядрами, не отличающимися по строению от ядра вегетативной формы. Незрелые цисты содержат 1-2 или 3 ядра. Имеют вакуоль сгликогеном. Часть цист хроматоидные тела.
С фекалиями цисты выбрасываются во внешнюю среду и при попадании в желудочно-кишечный тракт человека после метацистной стадии развития (деления на 8 дочерних амёб) образуют просветные формы.
Цисты могут сохранять жизнеспособность в воде и влажной почве более месяца[1].
кишечная амеба человека - Entamoeba coli. Однако среди обитающих в кишечнике человека амеб имеется один вид - дизентерийная амеба - Entamoeba histolytica , который может быть возбудителем тяжелой формы кишечного колита - амебиаза. Амеба эта имеет 20 - 30 мкм в диаметре, подвижна. Живет она в толстых кишках человека и обычно питается бактериями, не нанося никакого вредаНо в ряде случаев дизентерийная амеба начинает вести себя иначе: проникает под слизистую оболочку кишки, начинает там питаться и усиленно размножаться. Слизистая кишечника изъязвляется, результатом чего бывает тяжелый кровавый понос (колит). Распространение кишечных амеб осуществляется при помощи цист, выходящих вместе с фекальными массами наружу. Цисты очень стойки и длительное время сохраняют жизнеспособность и инвазионность (способность к заражению при попадании в кишечник человека). По строению цист можно установить вид амебы. Кишечная амеба Entamoeba coli имеет восьмиядерные цисты, тогда как дизентерийная (Entamoeba histolytica) - четырехъядерные. В цистах есть особые ярко окрашивающиеся включения - хроматоидные тельца. При сильном заражении с экскрементами выводится до 300 млн. цист в день. Кишечные амебы человека распространены по всему земному шару.
|
Амеба из кишечника человека (по Веньону). А - дизентерийная амеба (Entamoeba histolytica); Б - кишечная амеба (Entamoeba coll): 1 - ядро, 2 - заглоченные бактерии, 3 - заглоченные эритроциты
В кишечнике человека и ряда позвоночных обитает большое количество видов паразитических амеб, которые питаются содержимым кишечника, бактериями и большей частью не причиняют никакого вреда хозяину. Примером может служить кишечная амеба человека - Entamoeba coli ( рис. 4 ). Однако среди обитающих в кишечнике человека амеб имеется один вид - дизентерийная амеба - Entamoeba histolytica , который может быть возбудителем тяжелой формы кишечного колита - амебиаза. Амеба эта имеет 20 - 30 мкм в диаметре, подвижна ( рис. 4 , рис. 5 ). Живет она в толстых кишках человека и обычно питается бактериями, не нанося никакого вреда.
Подобное явление, когда патогенный паразитический организм не проявляет своей патогенности, называется носительством.
Но в ряде случаев дизентерийная амеба начинает вести себя иначе: проникает под слизистую оболочку кишки, начинает там питаться и усиленно размножаться. Слизистая кишечника изъязвляется, результатом чего бывает тяжелый кровавый понос (колит).
Распространение кишечных амеб осуществляется при помощи цист, выходящих вместе с фекальными массами наружу. Цисты очень стойки и длительное время сохраняют жизнеспособность и инвазионность (способность к заражению при попадании в кишечник человека). По строению цист можно установить вид амебы.
Кишечная амеба Entamoeba coli ( рис. 5 ) имеет восьмиядерные цисты, тогда как дизентерийная (Entamoeba histolytica) - четырехъядерные. В цистах есть особые ярко окрашивающиеся включения - хроматоидные тельца. При сильном заражении с экскрементами выводится до 300 млн. цист в день. Кишечные амебы человека распространены по всему земному шару.
Ротова́я амё́ба (лат. Entamoeba gingivalis) — это представитель подцарства простейшие (Protozoa), который встречается в кариозных зубах более чем у 25 % здоровых людей.
Патогенного действия не вызывает (т.е. не является паразитом), хотя обнаруживается при амфодонтозе, гайморите и остеомиелите челюстей.
Локализируется в альвеолах зубов, белом зубном налете и в криптах нёбных миндалин.
Существует только в форме трофозоита (вегетативная форма). Размеры колеблются от 6 до 60 мкм. Цитоплазма четко разделена на два слоя, где можно рассмотреть фагоцитированныебактерии и лейкоциты на разных стадиях пищеварения, при кровотечении из дёсен ротовая амёба может захватывать и эритроциты. Цист не образует. Ядро живой амёбы не визуализируется. Движение медленное, ложноножки широкие.
Нередко выделяется в мазках из десневых карманов при воспалительно-дистрофической форме пародонтита.[1]
Путь передачи — алиментарный. Заражение происходит через грязную воду, немытые овощи и фрукты.
Диагностика — обнаружение в соскобе кариозных зубов и мокроте трофозоитов.
Профилактика включает соблюдение гигиены полости рта, кипячение воды, мытье фруктов и овощей перед употреблением.
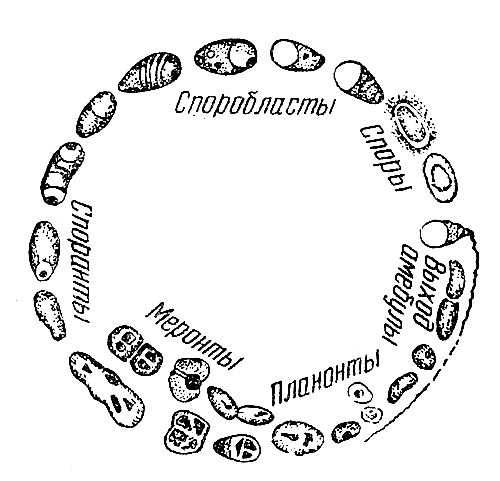

Вопрос №10
Эти простейшие передвигаются при помощи жгутиков, чем и объясняется название класса. Чаще всего имеется один жгутик, расположенный на переднем конце тела, реже - два, несколько или даже много жгутиков, которые отходят в разных местах поверхности тела. Оболочка тела жгутиковых, как правило, плотная, что придает определенную форму телу и облегчает быстрое передвижение. Есть жгутиковые, у которых оболочка очень тонка и они могут медленно передвигаться, переливаясь всем телом во временные выросты тела - ложноножки, или псевдоподии. такие формы наряду с ложноножками имеют и жгутики. Возможно, что древние жгутиковые имели одновременно и просто устроенные жгуты и псевдоподии. Жгутики, создавая водоворот, во многих случаях также способствуют поступлению пищи в цитоплазму. Многие представители описываемого класса имеют наружные скелеты из клетчатки или других веществ. У форм, постоянно находящихся в толще воды, скелет имеет нередко длинные выросты, благодаря чему увеличивается поверхность тела, что замедляет погружение таких организмов и облегчает их парение в воде.
Питание жгутиковых представляет большой интерес. В этом классе имеются как растительные, так и животные формы, а также миксотрофы, которые могут питаться и как автотрофы, и как гетеротрофы. Таким образом, в классе жгутиковых обе главные ветви органического мира - растения и животные - еще тесно связаны между собой.
Хлорофилл у автотрофных жгутиковых находится в пластидах, или хроматофорах. Нередко присутствие хлорофилла маскируется пигментами других цветов: желтого, красного и т. д. при переходе от автотрофного питания к гетеротрофному хроматофоры постепенно уменьшаются и, наконец, совсем исчезают. Автотрофные жгутиковые часто имеют особые глазки с пигментом, у многих - красным. Благодаря глазкам эти простейшие отыскивают освещенные места, где в их клетках может совершаться фотосинтез.
Жгутиковые всегда делятся продольно - вдоль длиной оси тела. У многих форм половой процесс чередуется с бесполым размножением. У целого ряда жгутиковых дочерние особи после деления не расходятся, а остаются вместе, вследствие чего образуются колонии. Форма колоний различная: у сидячих форм древовидная, у плавающих - шаровидная (например, у часто встречающегося в прудах вольвокса) или другая. Количество клеток в колонии у разных видов различно: от 2-4 особей до нескольких тысяч.
Жгутиковые обитают в морях и разнообразных пресных водоемах (от маленьких луж до больших озер), многие представители этого класса - паразиты. В стоячих пресных водоемах, в особенности небольших (лужах, прудах), некоторые виды жгутиковых развиваются массами, и вода становится зеленой или другой окраски - в зависимости от цвета хроматофоров. Это явление (которое вызывается также и различными низшими водорослями) носит название цветения: оно может быть очень вредным.
Разные виды паразитических жгутиковых у человека обитают в различных органах. Циклы их развития очень разнообразны.
Лейшмании (от имени У. Лейшмана) — род паразитических простейших, вызывающих лейшманиозы[4]. Переносчиками лейшманий являются москиты из рода Phlebotomus в Старом Свете, и из рода Lutzomyia в Новом Свете. Естественным резервуаром разных видов служат позвоночные животные, относящиеся к шести отрядам млекопитающих и ящерицам[5]; впрочем, на основании изоферментного анализа паразитов ящериц было предложено выделить в отдельный род отряда трипаносоматид — Sauroleishmania[6]. Лейшмании в основном поражают грызунов, собак и людей, но также отмечены случаи поражения у даманов, кошек и лошадей. В Новом Свете инфекция встречается у опоссумов, ленивцев и броненосцев[7]; зарегистрированы случаи лейшманиоза у кенгуру в Австралии[8]. По оценкам ВОЗ от начала 1990-х, лейшманиозами больны около 12 миллионов человек в 88 странах.
Содержание [убрать] 1 Происхождение 2 Морфология 3 Жизненный цикл 4 История открытия 5 Классификация 6 Геномика 7 Литература |
[править]Происхождение
Происхождение лейшманий неясно[9][10]. Одна из теорий предполагает африканское происхождение, с последующей миграцией в Северную и Южную Америку. Другая — миграцию из Северной и Южной Америки через Берингов перешеек, около 15 миллионов лет назад. Третья предлагает палеарктическое происхождение[11].
[править]Морфология
Амастиготы Leishmania tropica, расположенные внутриклеточно в макрофагах.

АмастиготыLeishmania mexicana.
Лейшмании существуют в двух морфологических формах — промастиготы (с длинным передним жгутиком, веретенообразные, удлинённые, подвижные)[12] в насекомом-хозяине и на искусственных питательных средах, и амастиготы (с коротким жгутиком, круглые или овальные, неподвижные, расположенные внутриклеточно) в организме позвоночных.
Промастиготы разных видов практически неразличимы, но амастиготы некоторых видов возможно предположительно опознать по некоторым внешним признакам либо по характерному распределению внутри макрофагов. Так, амастиготы L. major обычно крупнее (4—5 микрон), чем L. tropica (1,5—2,5 микрон; оба эти вида сосуществуют в некоторых очагах), а число их внутри каждой клетки меньше: L. major, как правило, не больше десяти промастигот на макрофаг (обычно 2—4 амастиготы), в то время как L. tropica — до ста паразитов на клетку[13]. Leishmania mexicana часто располагается в крупных вакуолях по периферии клетки, формируя характерные «гирлянды»[14].
[править]Жизненный цикл
Лейшмании, как и все трипаносоматиды, являются облигатными паразитами. Жизненный цикл лейшманий включает двух хозяев: млекопитающее и насекомое (москит).
Москиты заражаются лейшманиями, когда пьют кровь заражённого млекопитающего. Кровью питаются только самки москитов. Лейшмании вместе с проглатываемой кровью проникают в пищеварительный канал москита. В задней части средней кишки вокруг проглоченной крови у москита (как и у некоторых других кровососущих насекомых, например, комаров) образуется так называемая перитрофическая матрица.
Промастиготы лейшманий размножаются в пищеварительном канале самок москитов. Приблизительно через неделю инфекция распространяется до верхних отделов пищеварительного канала и паразиты блокируют просвет канала своими телами и секретируемым им гелем. Когда самка кусает потенциального хозяина, она выделяет в кожу свою слюну. Самка с блокированным пищеварительным каналом не может глотать, и у неё возникают спастические движения, в результате которых она отрыгивает промастиготы в ранку на коже хозяина.
В среднем во время укуса инфицированным москитом в кожу попадает 102 — 103 промастигот.
Первыми на место повреждения прибывают полиморфоядерные нейтрофилы, которые фагоциритуют паразитов. Внутри нейтрофилов лейшмании не размножаются и не превращаются в амастиготы. Затем, когда нейтрофилы переходят в фазу апоптоза, они уничтожаютсямакрофагами, и лейшмании проникают в макрофаги, не вызывая иммунного ответа.[15] Макрофаги являются основными клетками-хозяевами лейшманий в организме млекопитающих. Внутри макрофагов лейшмании трансформируются во внутриклеточную морфологическую форму — амастиготы.
Внутри макрофага лейшмании заключены в так называемые "паразитифорные вакуоли", которые образуются от слияния первичной фагосомыc лизосомами и эндосомами. В них промастиготы транформируются в амастиготы. При этом происходят изменения в морфологии — продолговатые промастиготы с длинным жгутиком превращаются в овальные амастиготы с коротким жгутиком, в метаболизме, чтобы приспособиться к кислой среде, и в биохимическом составе мембраны. Трансформация занимает от двух до пяти дней. Амастиготы способны выживать в кислой среде этих вакуолей и питаться их содержимым. Внутри вакуоли амастиготы медленно размножаются, и каждый цикл размножения занимает около 24 часов.
При кожном лейшманиозе инфильтрат образуется в коже и содержит преимущественно макрофаги, а также лимфоидные клетки и немного плазматических клеток. При висцеральном лейшманиозе очаги инфекции образуются в органах ретикуло-эндотелиальной системы.[16]
[править]История открытия
Факсимиле иллюстрации со страницы 935 из статьи Боровского «О сартовской язве». Рисунки поменяны местами: первый на месте второго, и наоборот.
Первое точное описание лейшмании в срезах кожных язв было опубликовано российским хирургом П. Ф. Боровским в «Военно-медицинском журнале» в 1898 году в статье «О сартовской язве». Боровский верно описал структуру паразитов и их отношение к тканям хозяина. Наблюдения Боровского подтвердил его коллега по Ташкентскому военному госпиталю К. Шульгин в журнале «Русский врач» в 1902 г. В 1903 г. американский врач Райтопубликовал независимое описание паразитов в кожной язве у девочки из Армении, которое принципиально не отличалось от описания Боровского, но содержало качественные иллюстрации. Райт предложил для открытого организма название Helcosoma tropicum. По-видимому, Райт не знал о работе Боровского, а также о появившемся в том же году описании паразитов, вызывающих кала-азар, или не связал описание возбудителя кала-азара со своим[17].
В 1904 г. Марциновский и Богров опубликовали на русском и немецком описание возбудителя восточной язвы, снабжённое микрофотографиями, для которого они предложили название Ovoplasma orientale, причём, по всей видимости, работали они, не зная о работах Райта, хотя были знакомы с публикацией Боровского. В упомянутой статье они преуменьшают заслуги Боровского и неверно приводят его наблюдения. Хотя наблюдения Марциноского и Богрова опубликованы через несколько месяцев после Райта, их исследование было проведено на несколько месяцев раньше[17].
[править]Классификация
В классификации лейшманий можно выделить четыре перекрывающихся периода. В начале XX в. основой классификации служили клинико-эпидемиологические и морфологические признаки, и лейшмании подразделялись на возбудителей висцерального лейшманиоза — L. donovani и возбудителей кожного лейшманиоза — L. tropica.
В 1913—1915 году В. Л. Якимов выделил два морфологических варианта возбудителя кожного лейшманиоза в Туркестане — с крупными или мелкими амастиготами — Leishmania tropica var. major и L. tropica var. minor[18], впоследствии они чаще обозначались как подвиды, без обозначения «var.». В 1940-х годах Кожевников и Латышев связали эти морфологические разновидности с двумя разными формами кожного лейшманиоза — антропонозным (L. tropica) и зоонозным (L. major)[19]. Впоследствии эти два подвида было предложено считать отдельными видами — L. tropica и L. major на основании клинико-эпидемиологических особенностей[20][21]. Морфологическая классификация применяется на практике и в начале XXI века; так, в провинции Балхна севере Афганистана, где встречается инфекция обоими этими видами, вид лейшманий определяют микроскопически на основании размера и количества амастигот в мазках из язв[13].
Тенденция считать возбудителей кожного лейшманиоза подвидами L. tropica просуществовала довольно долго. Так, в 1953 году Biagi предложил назвать описанного им возбудителя кожного лейшманиоза в Центральной Америке L. tropica var. mexicana[14].
Серологические методы классификации лейшманий начали применяться с середины 1910-х годов. Они позволяли отличать виды или комплексы видов. Реакция агглютинации и реакция связывания комплемента показали, что L. donovani и L. infantum серологически неразличимы. В 1930-х годах использование антисыворток позволило различить эти два вида[22]. В дальнейшем, с применением моноклональных антител, стало возможно различать отдельные серотипы внутри видов[23].
Характеризация лейшманий методом электрофореза изоферментов разрабатывалась с 1970-х годов начиная с отдельных ферментов[24], затем комбинаций ферментов[25][26]. Для обеспечения достаточной разрешающей способности требуется применять системы из многих ферментов — десяти до пятнадцати[27]. Штаммы организмов с одинаковыми электрофоретическими профилями называеются «зимодем».
Наиболее полный банк лейшманий, охарактеризованных изоферментным анализом, хранится в университете Монпелье 1 (фр.). Харктеризация проводится на основании электрофореза 15 изоферментов с 1981 года. Начиная с 1989 года применяется метод изоэлектрического фокусирования[28], который имеет более высокую разрешающую способность, но применим только к шести ферментам[29].
Изоферментный анализ в последние четверть века является стандартным методом типирования лейшманий на видовом и подвидовом уровнях, потому что он был применён к наибольшему количеству самых разнообразных штаммов по сравнению с другими методами[30].
Зимодемы в лаборатории Монпелье получают обозначение, состоящее из сокращения MON (Монпелье), за которым следует порядковый номер зимодема. По состоянию на 2008 год, в лаборатории хранится более 3000 образцов лейшманий, разделённых на более чем 260 зимодемов.
Генетические методы, основанные на ПЦР, начали применяться с 1970-х годов. Эти методы имеют некоторые ограничения. В частности, изучение гетерозиготности и рекомбинации недостаточно разработаны для видов, в жизненном цикле которых нет гаплоидной стадии[30].
Разные методы классификации в основном совпадают в определении комплексов видов и генетических связей между ними.
На основании хода инфекции в моските-переносчике, род Leishmania делится на два подрода: Leishmania (Leishmania) и Leishmania (Viannia). Представители подрода Leishmaniaприкрепляются к эпителию средней кишки москита, а подрода Viannia — также к эпителию задней кишки. Представители подрода Leishmania распространены в Старом и Новом Свете, а подрода Viannia — только в Новом Свете[6].
Паразитов рептилий одни авторы выделяют в третий подрод лейшманий — Leishmania (Sauroleishmania), другие рассматривают его как отдельный род трипаносоматид — Sauroleishmania[6].
[править]Геномика
Геномы трёх видов (L. major, L. infantum и L. braziliensis) были отсеквенированы, при этом было выявлено более 8300 генов белков и 900 генов РНК. Почти 40% белок-кодирующих генов были распределены по 662 семействам, содержащим от 2 до 500 генов. Небольшие семейства в основном представлены генами, образующими тандемные повторы в различных локусах по всему геному. Каждая из 35 или 36 хромосом организована в небольшое число кластеров из 10-100 генов, расположенных на одной цепи ДНК. Эти кластеры могут быть расположены «голова к голове» (расходящиеся) или хвост к хвосту (сходящиеся). В последнем случае они могут быть разделены генами рРНК, тРНК или мяРНК. Транскрипция белок-кодирущих генов начинается в области расхождения кластеров и продолжается полицистронно до области переключения цепей между сходящимися кластерами. Теломеры лейшманий обычно относительно короткие и состоят из нескольких типов повторяющихся последовательностей[31].
В 2009 году было показано, что в организме насекомого у лейшманий происходит процесс обмена генетической информацией, предполагающий мейоз[32].
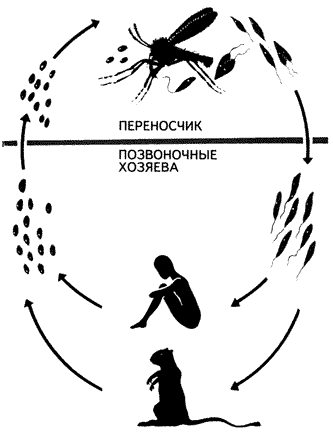
№ 99 Систематика, морфонология и биология возбудителей лейшманиозов. Лабораторная диагностика, профилактика, патогенное действие.
Лейшмании Leischmania (кл. Жгутиковые) — возбудители лейшманиозов. Заболевания человека вызываются несколькими видами и подвидами паразитов, которые объединяются в четыре комплекса: L. donovani — возбудитель висцерального лейшманиоза, L. tropica — возбудитель кожного лейшманиоза, L. mexicana — возбудитель лейшманиоза Центральной Америки, L brasiliensis — возбудитель бразильского лейшманиоза. Все виды сходны морфологически и имеют одинаковые циклы развития. Они существуют в двух формах: в безжгутиковой, или лейшманиальной, и жгутиковой, или промасти-готной (рис. 19.8).
Лейшманиальная форма очень мелка — 3—5 мкм в диаметре. Характерной чертой ее является круглое ядро, занимающее около 1/4 цитоплазмы; жгутика нет, но перпендикулярно клеточной поверхности располагается палочковидный кинетопласт. Эти формы обитают в клетках ретикулоэндотелиальной системы человека и ряда млекопитающих (грызунов, собак, лис). Промастиготная форма удлинена — до 25 мкм, спереди находится жгутик, у основания которого хорошо виден такой же кинетопласт, что и в безжгутиковой стадии паразита. Обитает в пищеварительной системе москитов. Безжгутиковая форма, посеянная на культуральную среду, превращается в жгутиковую.
Лейшманиозы широко распространены в странах с тропическим и субтропическим климатом на всех континентах там, где обитают москиты. Они - типичные природно-очаговые заболевания (см. § 18.13). Природными резервуарами являются грызуны, дикие и домашние хищники. Заражение человека происходит при укусе инвазированными москитами.
По патогенному действию лейшманий заболевания, которые они вызывают, делят на три основные формы: кожный, слизисто-кожный и висцеральный лейшманиозы.
Лабораторная диагностика основана на микроскопировании мазков из кожных язв при кожном и слизисто-кожном лейшманиозах, пунктатов лимфатических узлов и костного мозга при висцеральном лейшманиозе. В окрашенных препаратах обнаруживается лейшманиальная форма паразитов как внутри клеток, так и внеклеточно. В сомнительных случаях производят посев материала, взятого от больного, на специальную культуральную среду, на которой лейшмании приобретают промастиготную форму, активно передвигаются и легко обнаруживаются при микроскопировании. Используют также и биологические пробы — заражение лабораторных грызунов.
Профилактика — в первую очередь, это борьба с переносчиками и уничтожение природных резервуаров (грызунов и бродячих собак), а также профилактические прививки.
№ 100 Трипаносомы. Систематика, морфология, цикл развития, пути заражения, лабораторная диагностика, профилактика, патогенное действие.
Ttypanosoma brucei gambiense и T.b. rhodesiense (кл. Жгутиковые) — возбудители африканского трипаносомоза, или сонной болезни. Паразит имеет извилистую заостренную с обеих сторон форму. Длина его 17—28 мкм. Стадии, паразитирующие у человека, имеют один жгутик, ундулирующую мембрану сбоку и хорошо заметный кинетопласт у основания жгутика.
Трипаносомы поселяются у человека в крови, лимфе, спинномозговой жидкости, в тканях головного и спинного мозга и в серозных полостях. T.b. gambiense встречается в Западной Африке, а T.b. rhodesiense — в Восточной и Юго-Восточной Африке.
Жизненный цикл этих паразитов протекает в организме человека, домашних и диких млекопитающих, в первую очередь копытных. T.b. gambiense чаще поражает человека, свиней и собак, T.b. rhodesiense — диких животных — антилоп и носорогов. Переносчиком первого подвида является муха це-це Glossina palpalis, живущая поблизости от жилища человека, второго — G. morsitans, обитающая в открытых саваннах и саванновых лесах. В связи с этим сонная болезнь, возбудителем которой является T.b. gambiense, встречается в антропогенных очагах культурных ландшафтов. Ежегодно регистрируется около 10 000 новых случаев заражения. Восточноафриканский трипаносомоз распространен значительно реже в естественной природе. В основном заболевают охотники, туристы, сезонные рабочие, каждый год — около 1500 человек.
Лабораторная диагностика — исследование мазков крови и спинномозговой жидкости больного для выявления в них возбудителя. Используются также иммунологические реакции и заражение лабораторных животных.
Профилактика — кроме борьбы с переносчиками применяют профилактическое лечение здоровых людей, живущих в очагах трипаносомоза, делающее организм невосприимчивым к инвазии. Практиковавшийся ранее отстрел диких животных, являющихся природным резервуаром паразита, вряд ли рационален в связи с возможным нарушением экологического баланса, который складывался в биогеоценозах на протяжении тысячелетий.
Диагностика — в острой форме заболевания возможно обнаружение трипаносом в крови. При хроническом течении рационально введение крови больного морским свинкам, у которых они обнаруживаются в большом количестве на 14-е сутки. Существует еще и своеобразный метод диагностики — кормление на больном неинвазированных переносчиков-клопов, в кишечнике которых трипаносомы быстро размножаются и легко обнаруживаются. Применяют также и методы иммунодиагностики.
Трипаносомы представляют большой интерес не только потому, что способны вызывать у человека серьезные, смертельно опасные заболевания. Колоссальная экологическая пластичность, обеспечивающая им эффективное паразитирование в организме хозяина на протяжении нескольких лет, в условиях постоянно действующих механизмов иммунитета, позволяет этим паразитам осваивать и новых хозяев.
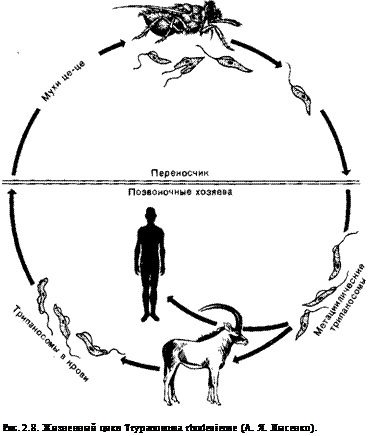
№ 101 Трихомонада. Систематика, морфология, цикл развития, пути заражения, лабораторная диагностика, профилактика, патогенное действие.
ТРИХОМОНАДА ВЛАГАЛИЩНАЯ - Trichomonas urogenitalis (vaginalis) - возбудитель мочеполового трихомониаза - антропоноза.
Географическое распространение - повсеместно.
Локализация - у женщин паразитирует во влагалище, в цервикальном канале, уретре, мочевом пузыре и бартолиниевых железах. У мужчин - в уретре, мочевом пузыре, предстательной железе и куперовых железах.
Морфологическая характеристика. Существует лишь в вегетативной форме.
Вегетативная форма размером 14 - 30 мкм, грушевидная, имеет аксостиль, 1 ядро, ундулирующую мембрану, которая достигает только середины тела. Есть 3 - 4 свободных жгутика и один (пятый) тянется вдоль ундулирующей мембраны. Цитостом отсутствует.
Цикл развития.
Инвазионная форма. Вегетативная форма трихомонады попадает в организм человека в основном половым путем, изредка бытовым (через общее полотенце, мочалку и т. д.). Паразит во внешней среде не устойчив и гибнет уже при температуре 39 - 40°С.
Патогенное действие. В слизистых оболочках пораженных органов возникает воспалительный процесс. Возможно скрытое течение заболевания со слабо выраженными симптомами (носительство).
Источник заражения - больной человек и носитель.
Диагностика. Микроскопическое исследование свежих мазков из влагалища и уретры (окраска метиленовым синим или по Романовскому - Гимза).
Профилактика: а) общественная - активное выявление больных и носителей, их лечение; б) личная - соблюдение гигиенических норм в банях, бассейнах, душах и так далее.
ТРИХОМОНАДА КИШЕЧНАЯ - Trichomonas hominis (intestinalis) - возбудитель кишечного трихомониаза - антропоноза.
Географическое распространение - повсеместно.
Локализация - паразит в просвете слепой кишки и нижних отделах тонкого кишечника.
Морфологическая характеристика. Существует лишь в вегетативной форме. Вегетативная форма размером 8 -12 мкм, грушевидная, имеет аксостиль, цитостом, 1 ядро, 3-4свободных жгутика и один жгутик тянется вдоль всего тела по краю ундулирующей мембраны.
Цикл развития.
Инвазионная форма. Вегетативная форма попадает к человеку через рот. Заражение происходит при проглатывании трихомонады с немытыми овощами, фруктами, некипяченой водой, то есть фекально - оральным способом.
Патогенное действие. Вопрос о патогенности спорный. Считается, что трихомонада может вызывать воспалительный процесс в кишечнике, так как иногда обнаруживают трихомонад с заглоченными эритроцитами. Трихомониаз чаще сопутствует патологическим процессам в кишечнике, вызванным другими причинами.
Источник заражения - больной человек и носитель.
Диагностика. Микроскопическое исследование свежих мазков жидких фекалий. Обнаружение в них вегетативных форм.
Профилактика: а) общественная - охрана воды и почвы от загрязнения фекалиями; б) личная - соблюдение правил личной гигиены (мытье рук, овощей, фруктов и кипячение воды).
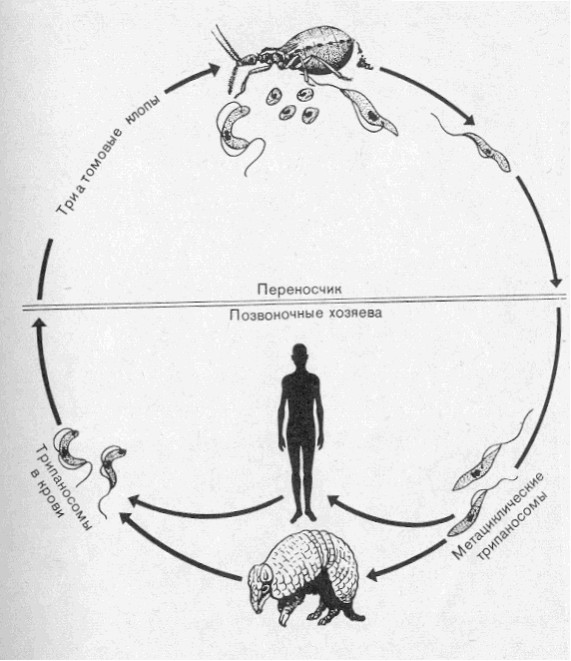
№ 97 Лямблия. Морфология, цикл развития. Пути заражения. Диагностика, профилактика.
В тонкой кишке человека паразитирует единственный вид простейших — лямблия Lamblia intestinalis (кл. Жгутиковые) — возбудитель лямблиоза, которым чаще болеют дети. Форма паразита напоминает грушу, разрезанную вдоль. Длина тела 10—18 мкм. В расширенной части, на уплощенной стороне расположен присасывательный диск, с помощью которого лямблии присасываются к ворсинкам кишечника. Вдоль тела проходят две тонкие опорные органеллы — аксостили. Симметрично в клетке располагаются два ядра и четыре пары жгутиков
Трофозоиты используют питательные вещества с поверхности клеток кишечного эпителия. Захват пищи осуществляется пиноцитозом.
Большие количества лямблий, которые покрывают обширные поверхности кишечной стенки, нарушают процессы всасывания и пристеночного пищеварения. Попадая в нижние отделы тонкой кишки, лямблии инцистируются. Зрелые цисты имеют овальную форму, четыре ядра и несколько аксостилей. Во внешней среде цисты сохраняют жизнеспособность в течение нескольких недель. Заражение человека происходит при проглатывании цист.
Лабораторная диагностика — обнаружение цист в фекалиях и тро-фозоитов в содержимом двенадцатиперстной кишки, полученном при дуоденальном зондировании.
Личная профилактика — соблюдение правил гигиены питания. Общественная профилактика — санитарное благоустройство туалетов, предприятий общественного питания.
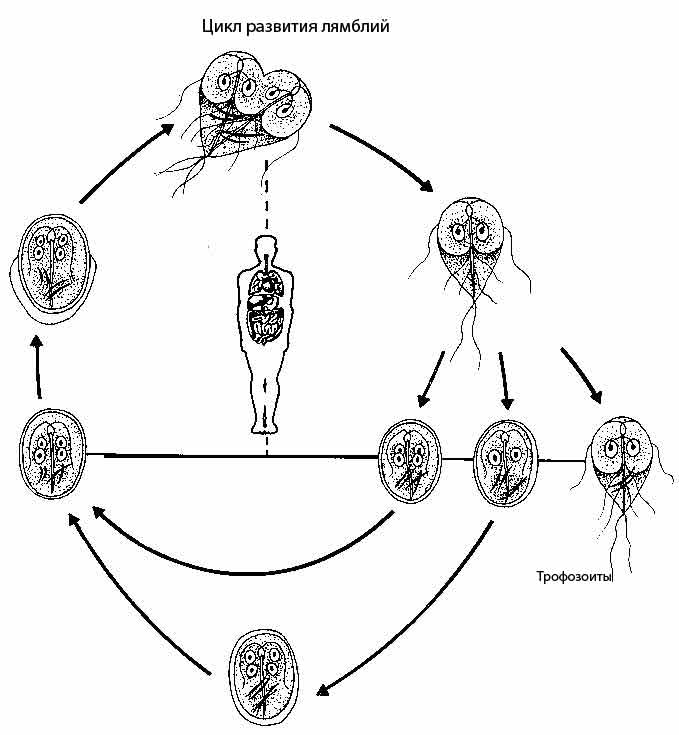
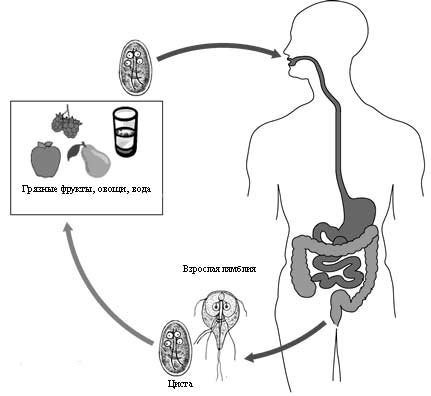
Вопрос № 11
Класс Инфузории
Это наиболее сложно устроенные простейшие. Характерными особенностями организации инфузорий являются: движение с помощью ресничек, наличие двух ядер— большого и малого — с разными функциями и полового процесса—конъюгации.
Инфузория-туфелька — обитатель мелких стоячих водоемов. Ее длина достигает 0,1—0,3 мм. Покрыта она пелликулой, поэтому форма тела постоянная и напоминает изящную дамскую туфельку, откуда и ее название (рис. 10.3).
Движение туфельки осуществляется с помощью многочисленных (более 10 тыс.) ресничек, расположенных правильным продольными рядами. Они совершают согласованные волнообразные колебания.
Питание происходит следующим образом. На одной из сторон тела туфельки имеется воронкообразное углубление, ведущее в рот и трубчатую глотку. С помощью ресничек, выстилающих воронку, пищевые частицы (бактерии, одноклеточные водоросли, детрит) загоняются в рот, а затем в глотку. Из глотки пища путем фагоцитоза проникает в цитоплазму Образовавшаяся при этом пищеварительная вакуоль подхватывается круговым током цитоплазмы. В течение 1—1,5 ч пища переваривается, всасывается в цитоплазму, а непереваренные остатки через отверстие в пелликуле — порошицу — выводятся наружу. При благоприятных температурных и пищевых условиях за сутки туфелька способна потребить столько пищи, сколько весит сама.
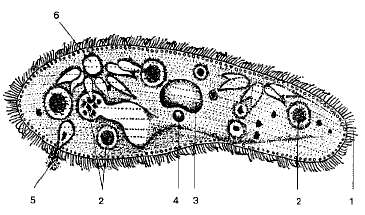
Рис. 10.3. Инфузория-туфелька: 1 —реснички; 2 — пищеварительные вакуоли; 3 — большое ядро (макронуклеус); 4 —малое ядро (микронуклеус); 5 — порошица; б — сократительная вакуоль.
Сократительных вакуолей в цитоплазме туфельки две: в передней и задней частях тела. Устроены они более сложно, чем у простейших других классов. Вода и конечные продукты жизнедеятельности из цитоплазмы сначала проникают в приводящие канальцы, а затем из них — в центральную вакуоль, откуда выводятся наружу. Сокращаются вакуоли попеременно каждые 20—30 с.
Ядерный аппарат инфузории-туфельки устроен сложно и представлен большим бобовидной формы полиплоидным ядром, или макронуклеусом, регулирующим вегетативные функции (питание, дыхание, выделение), и малым ядром, или микронуклеусом,играющим особую роль в половом процессе.
Размножение бесполое. Сначала делятся оба ядра: большое амитотически, а малое митотически, а затем происходит деление тела инфузории-туфельки пополам в поперечном направлении. Недостающие органеллы заново развиваются у обеих дочерних особей.
Бесполое размножение после ряда поколений сменяется периодически наступающим половым процессом — конъюгацией. При этом две инфузории прикладываются друг к другу сторонами, где расположен рот. Пелликула в месте контакта особей растворяется, и между ними образуется цитоплазматический мостик. Большое ядро разрушается и в половом процессе не принимает участия. Малые ядра делятся мейотически. Из четырех образовавшихся в каждой инфузории гаплоидных ядер три распадаются. Оставшееся четвертое ядро делится митотически еще один раз. Одно из двух образовавшихся ядер (стационарное) остается в той же инфузории, а другое (мигрирующее) переходит в клетку партнера по конъюгации. После слияния стационарного и мигрирующего ядер образуется диплоидное ядро с рекомбини-рованным генетическим материалом. В каждой из инфузорий диплоидное ядро делится несколько раз, и после ряда преобразований формируются малое и обновленное большое ядро. Через некоторое время инфузории приступают к активному бесполому размножению делением.
Эти простейшие покрыты как будто тончайшей «шерстью» — ресничками. У каждой инфузории их около 10—20 тыс. Реснички — это те же укороченные жгутики. Движение ресничек волнообразно, как колыхание пшеничного поля. Синхронно взмахивая около 30 раз в секунду, ряды ресничек, как вёсла множества гребцов, движут инфузорию вперёд. Этот «скоростной» способ передвижения, конечно, не сравнить с неспешным перетеканием амёбы. Помещённая в каплю воды под окуляр микроскопа, инфузория 601c часто столь стремительно пересекает поле зрения микроскопа, что напоминает маленькую ракету. За секунду инфузория часто проплывает 10—20 длин своего тела.
Инфузории — самые совершенные создания природы среди простейших. Это процветающая группа животных, к которой принадлежит более 7 тыс. видов.
Можно сказать, что эволюция, начав «лепку» тела простейших с бесформия амёбы, постепенно пришла к строго определённым формам тела у инфузорий.
У большинства инфузорий есть хорошо оформленная глотка — довольно глубокая впадина или воронка на теле. Бактерии и прочая добыча инфузорий, если они попали в глотку, уже никуда не денутся — взмахи ресничек отправят их на дно глотки, где расположен рот.
Другое важное усовершенствование строения инфузории — наличие в клетке не менее двух ядер. Зачем это нужно?
Ядро, как известно, является хранилищем «инструкций и чертежей», определяющих работу клетки. Два ядра можно сравнить с двумя залами библиотеки. В первом — книги (чертежи и инструкции) находятся в постоянном обороте, на руках, их читают, они часто получают повреждения. Это большое ядро инфузории (макронуклеус). Оно руководит всей повседневной деятельностью клетки. Во втором же зале нашей библиотеки хранятся точно такие же книги (копии тех же инструкций и чертежей), но в неприкосновенном виде. Здесь они сберегаются для будущих поколений. Это малое ядро инфузории (микронуклеус). Оно необходимо только в самые важные моменты жизни инфузорий — при половом процессе (о котором речь пойдёт ниже).
Половой процесс у инфузорий
Каждая инфузория регулярно делится надвое, порождая две свои точные копии, похожие, как близнецы. На первый взгляд может показаться, что инфузории сумели для себя осуществить давнюю мечту человека о бессмертии и вечной молодости. Но это не так. Если бесконечно выращивать потомство одной-единственной инфузории, в конце концов через несколько сот поколений станут заметны признаки вырождения, старения.
Персонажи некоторых сказок (например, герои «Конька-горбунка» Павла Ершова) пытались омолодиться, бросаясь в котёл с кипящей водой. Часто это приводило к самым печальным последствиям. Для инфузорий омоложение тоже удивительным образом переплетено со смертью. Особь умирает и в то же время — рождается заново. Каков же рецепт омоложения, «придуманный» инфузориями?
Различные инфузории, в том числе туфелька и трубач (вверху в центре и справа).
«Рождаются заново» инфузории в ходе полового процесса, именуемого конъюгацией.
При этом две особи прикладываются друг к другу брюшной стороной, где находится рот. Большие их ядра разрушаются, а малые начинают делиться. В конце концов у каждой особи образуется по два приблизительно одинаковых ядра. И тут наступает самый важный момент: инфузории обмениваются ядрами. Одно из ядер (мужское) инфузория дарит партнёру, второе (женское) оставляет себе. Затем у каждой особи мужские ядра сливаются с женскими.
Расходятся после этого уже совсем не те существа, которые встретились. Теперь каждое из них — наполовину прежняя особь, а наполовину — копия партнёра. Пожалуй, инфузории могут в самом точном и буквальном смысле назвать своих «супругов» «половиной» друг друга!
Известно около 6000 видов, относящихся к классу Инфузорий. Большинство представителей — это обитатели морских и пресных водоемов, некоторые обитают во влажной почве или песке. Многие виды являются паразитами человека и животных.