

Контроль:
1. Типы клеточного строения.
2. Компартментализация клетки.
3. Структурно-функциональная организация эукариотической клетки.
4. Молекулярное строение и функциональные компоненты клеточных мембран.
5. Типы и функции мембранных липидов.
6. Типы и функции мембранных белков.
7. Основные функции углеводов биомембран.
8. Избирательная проницаемость клеточных мембран.
9. Роль мембран в развитии патологии.
1)Среди живых организмов только вирусы не имеют клеточного строения. Все остальные организмы представлены клеточными формами жизни. Различают два типа клеточной организации: прокариотический и эукариотический. К прокариотам относят бактерии и цианобактерии (сине-зеленые), к эукариотам — растения, грибы и животных. Прокариотические клетки устроены сравнительно просто, они не имеют ядра. Область расположения ДНК в цитоплазме называется нуклеоид. Единственная молекула ДНК — кольцевая и не связана с белками; клетки меньше эукариотических, в состав клеточной стенки входит гликопептид — муреин; мембранные органоиды отсутствуют, их функции выполняют впячивания плазматической мембраны (мезосомы); рибосомы мелкие, микротрубочки отсутствуют, поэтому цитоплазма неподвижна, а реснички и жгутики имеют особую структуру. Эукариотические клетки имеют ядро, в котором находятся хромосомы — линейные молекулы ДНК, связанные с белками, в цитоплазме расположены различные мембранные органоиды. Растительные клетки отличаются наличием толстой целлюлозной клеточной стенки, пластида, крупной центральной вакуоли, смещающей ядро к периферии. Клеточный центр высших растений не содержит центриоли. Запасным углеводом является крахмал. Клетки грибов имеют клеточную стенку, содержащую хитин; в цитоплазме имеется центральная вакуоль, отсутствуют пластиды. Только у некоторых грибов в клеточном центре встречается центриоль. Главным резервным углеводом является гликоген. Животные клетки не имеют клеточной стенки, не содержат пластид и центральной вакуоли, для клеточного центра характерна центриоль. Запасным углеводом является гликоген. В зависимости от количества клеток, из которых состоят организмы, их делят на одноклеточные и многоклеточные. Одноклеточные организмы состоят из одной-единственной клетки, выполняющей функции целостного организма. Одноклеточными являются все прокариоты, а также простейшие, некоторые зеленые водоросли и грибы. Тело многоклеточных организмов состоит из множества клеток, объединенных в ткани, органы и системы органов. Клетки многоклеточного организма специализированы для выполнения определенной функции и могут существовать вне организма лишь в микросреде, близкой к физиологической (например, в условиях культуры тканей). Клетки в составе многоклеточного организма различаются по размерам, форме, структуре и выполняемым функциям. Несмотря на индивидуальные особенности все клетки построены по единому плану и имеют много общего. 2)Принцип компартментализации клеток эукариот постулирует, что биохимические процессы в клетке локализованы в определённых отсеках, покрытых оболочкой из бислоя липидов. Большинство органоидов в эукариотической клетке являются компартментами — митохондрии, хлоропласты, пероксисомы, лизосомы, эндоплазматический ретикулум, ядро клетки и аппарат Гольджи. Содержание 1 Функции 2 Классификация 3 Прокариоты 4 См. также 5 Примечания Функции Внутри компартментов, окруженных биослоем липидов, могут существовать различные значения pH, функционировать разные ферментативные системы. Принцип компартментализации позволяет клетке выполнять разные метаболические процессы одновременно. В цитозоле митохондрий находится окислительная среда, в которой NADH окисляется в NAD+. Квинтэссенцией(самое важное) принципа компартментализации можно считать аппарат Гольджи, в диктиосомах(составная часть комплекса Гольджи, система мембран, сложенных стопкой.) которого работают различные ферментативные системы, осуществляющие, например, разные стадии посттрансляционной модификации белков. Классификация[править | править вики-текст] Выделяют следующие клеточные компартменты: Ядро (внутреннее содержимое ядра) Пространство цистерн эндоплазматического ретикулума (переходящее в перинуклеарное пространство) Аппарат Гольджи Лизосомы Митохондрии (подразделяются на два компартмента — матрикс и межмембранное пространство) Хлоропласты (у высших растений подразделяются на три компартмента — межмембранное пространство, строму и внутреннюю полость тилакоидов) Цитозоль Прокариоты[править | править вики-текст] В любой клетке существует два генеральных микрокомпартмента, разделённые унитарной мембраной — цитоплазматический и экзоплазматический. Бактерии, обладающие грамотрицательным морфотипом, имеют ещё и третий генеральный микрокомпартмент — периплазматический, который расположен между цитоплазматической мембраной и наружной мембраной[1]. Иногда специализированный микрокомпартмент размещается сразу в нескольких генеральных компартментах, то есть имеет смешанную локализацию. Одним из примеров этого служит ундулиподия.
3)3)Наиболее сложная организация присуща эукариотическим клеткам животных и растений. Строение клеток животных и растений характеризуется принципиальным сходством, но форма, размеры и масса их чрезвычайно разнообразны и зависят от того, является ли организм одноклеточным или многоклеточным. Например, диа-томовые водоросли, эвгленовые, дрожжи, миксомицеты и простейшие являются одноклеточными эукариотами, тогда как организмы подавляющего большинства других типов являются многоклеточными эукариотами, количество клеток у которых составляет от нескольких (например, у некоторых гельминтов) до миллиардов (у млекопитающих) на организм. Организм человека состоит из около 10 различных клеток, которые различаются между собой по осуществляемым ими функциям.
В случае человека насчитывают более 200 типов разных клеток. Наиболее многочисленными клетками в организме человека являются эпителиальные клетки, среди которых различают орого-вевающие клетки (волос и ногтей), клетки, обладающие всасывательной и барьерной функциями (в желуд очно-кишечном тракте, мочеполовых путях, роговице, влагалище и других системах органов), клетки, выстилающие внутренние органы и полости (пневмо-циты, серозные клетки и многие другие). Различают клетки, обеспечивающие метаболизм и накопление резервных веществ (гепатоциты, жировые клетки). Большую группу составляют эпителиальные и соединительнотканные клетки, секретизирующие внеклеточный матрикс (амилобласты, фибробласты, остеобласты и другие) и гормоны, а также сократительные клетки (скелетных и сердечных мышц, радужной оболочки и других структур), клетки крови и иммунной системы (эритроциты, нейтрофилы, эозинофилы, базофилы, Т-лимфоциты и другие). Существуют также клетки, выполняющие роль сенсорных преобразователей (фоторецепторы, осязательные, слуховые, обонятельные, вкусовые и другие рецепторы). Значительное число клеток представлено нейронами и гли-альными клетками центральной нервной системы. Существуют также специализированные клетки хрусталика глаза, пигментные клетки и питающие клетки, далее следует назвать подовые клетки. Известны и многие другие типы клеток человека.
В природе не существует некой типичной клетки, ибо все они характеризуются чрезвычайным разнообразием. Тем не менее все эукариотические клетки существенно отличаются от прокариотических клеток по ряду свойств и прежде всего по объему, форме и размерам. Объем большинства эукариотических клеток превышает объем прокариотов в 1000-10 000 раз. Такой объем прокариотических клеток связан с содержанием в них различных органелл, осуществляющих всевозможные клеточные функции. Для эукариотических клеток характерно также наличие большого количества генетического материала, сосредоточенного в основном в относительно большом количестве хромосом, что обеспечивает им большие возможности в дифференцировке и специализации. Не менее важной особенностью эукариотических клеток является то, что им присуща компартментализация, обеспеченная наличием внутренних мембранных систем. В результате этого многие ферменты локализуются в определенных компартментах. Например, почти все ферменты, катализирующие синтез белков в животных клетках, локализованы в рибосомах, тогда как ферменты, катализирующие синтез фосфолипидов, в основном сосредоточены на клеточной ци-топлазматической мембране. В отличие от прокариотических клеток в эукариотических клетках имеется ядрышко.
Эукариотические клетки по сравнению с прокариотическими обладают более сложной системой восприятия веществ из окружающей среды, без чего невозможна их жизнь. Существуют и другие различия между эукариотическими и прокариотическими клетками.
Форма клеток бывает самой разнообразной и часто зависит также от выполняемых ими функций. Например, многие простейшие имеют овальную форму, тогда как эритроциты являются овальными дисками, а мышечные клетки млекопитающих вытянуты. Размеры эукариотических клеток являются микроскопическими (табл. 3).
Некоторые виды клеток характеризуются значительными размерами. Например, размеры нервных клеток у крупных животных достигают нескольких метров в длину, а у человека — до 1 метра. Клетки отдельных тканей растений достигают нескольких миллиметров в длину.
Считают, что чем крупнее организм в пределах вида, тем крупнее его клетки. Однако для родственных видов животных, различающихся по размерам, характерны и сходные по размерам клетки. Например, у всех млекопитающих сходны по размерам эритроциты.
Клетки различаются также и по массе. Например, одиночная клетка печени (гепатоцит) человека весит 19-9 г.
Соматическая клетка человека (типичная эукариотическая клетка) представляет собой образование, состоящее из множества структурных компонентов микроскопических и субмикроскопических размеров(рис. 46).
Использование электронной микроскопии и других методов позволило установить чрезвычайное разнообразие в структуре как оболочки и цитоплазмы, так и ядра. В частности, был установлен мембранный принцип строения внутриклеточных структур
Мембранная система. Эта система представлена клеточной плазматической (цитоплазматической) мембраной, цитоплазматической (эндоплазматической) сетью (рети-кулом) и пластинчатым комплексом Гольджи.
а) Плазматическая (цитоплаз-матическая) мембрана имеет толщину 8-12 нм и состоит из трех слоев, два из которых являются белковыми слоями толщиной по 3 нм каждый, а третий (внутренний) — двойным фосфолипидным слоем толщиной 6 нм (рис. 47). Плазматическая мембрана является полупроницаемой структурой. Через нее в клетку входят питательные вещества и выходят все «отходы» (продукты секреции). Она создает барьер проницаемости. В результате этого плазматическая мембрана регулирует обмен различными веществами между клеткой и внешней средой. В плазматической мембране содержатся многие важные ферменты, системы активного транспорта ионов натрия и калия при помощи АТФазы, а также системы транспорта аминокислот.
На поверхности плазматической мембраны обнаруживают ряд специальных образований в виде микроворсинок и ресничек. Микроворсинки очень часты в эпителии кишечника и почек. Реснички — это своеобразные выросты цитоплазмы. У эритроцитов мембрана является гладкой (элементарная мембрана). У некоторых одноклеточных организмов-эукариотов плазматическая мембрана также содержит реснички (микроворсинки), различные выпячивания, впячивания и выросты, переходящие в пузырьки. На внешней поверхности клеток животных обнаружены гликопротеиды как компоненты плазматической мембраны.
Предполагают, что поверхностные гликопротеиды обеспечивают адгезионную способность клеток в тканях, и, следовательно, слипание однотипных клеток. В мембранах эритроцитов содержится гликопротеид, получивший название гликофорина (м. м. 30 000). Этот гликопротеид состоит из 130 аминокислотных остатков и большого количества (60% всей молекулы) остатков сахара. Кроме того, в эритроцитарной мембране содержится белок спектрин, молекулы которого формируют скелет мембраны.
У клеток растений наружной структурой служит жесткая клеточная стенка, построенная из молекул целлюлозы, создающих очень прочные волокна, погруженные в матрикс из других поли-сахаридов и полимерного вещества лигнина. Клетки наружних слоев растений иногда покрыты очень тонким слоем восковидно-го вещества. На поверхности плазматических мембран имеются электрически заряженные группы, из-за которых поддерживается разность электрических потенциалов на мембранах. На поверхности плазматических мембран имеются также специфические рецепторы (участки распознания) для гормонов и других соединений. Кроме того, здесь же локализованы особые рецепторы, ответственные за индивидуальную тканевую совместимость. Считают, что рецепторные участки формируются гликопротеидами и ганглиозидами.
б) Цитоплазматическая (эндоплазматическая) сеть (рети-кулум) представлена пронизывающими однослойными мембранными полостями (трубочками, цистернами, вакуолями) разных размеров, заполненными белковыми гранулами.
Открыта К. Портером в 1945 г. Толщина трубочек и других структур этой сети равна 5-6 нм.
Различают гранулярный (шероховатый) эндоплазматический ретикулум, который выстлан множеством рибосом диаметром порядка 21-25 нм и молекулярной массой 4 х 106, служащих центрами синтеза молекул белков, и агранулярный (гладкий) эндоплазматический ретикулум, на котором нет рибосом, но на котором синтезируются липиды и углеводы. Степень насыщенности гранулярной эндоплазмати-ческой сети рибосомами определяет степень интенсивности синтеза белков. У человека и животных агранулярной сетью богаты клетки коры надпочечников, яичников и семенников, печени, скелетных мышц.
Цитоплазматическая сеть без перерыва соединена с цитоплаз-матической мембраной, ядерной мембраной и пластинчатым комплексом Гольджи. Это позволяет синтезируемым белкам проходить в комплекс Гольджи, откуда после специальной обработки они выводятся из клетки или идут на построение лизосом.
Плазматическая мембрана, мембрана эндоплазматической сети, а также ядер, митохондрий и хлоропластов (см. ниже) представляют собой чрезвычайно сложные структуры, обладающие рядом важ-.нейших биологических свойств. Многие мембраны содержат ферменты, транспортные системы, с помощью которых осуществляется перенос молекул питательных веществ и неорганических ионов внутрь клеток и внутри клеток, а также вывод из клеток продуктов жизнедеятельности. Мембранные структуры способны к самовосстановлению, если в них по каким-то причинам возникают повреждения.
В процессе искусственного растирания или гомогенизации клеток с экспериментальными целями образуются мелкие частицы диаметром 50—150 нм, состоящие из фрагментов эндоплазматическо-го ретикулума и плазматической мембраны. Эти структуры получили название микросом и их широко используют в лабораторной работе для решения тех или иных вопросов молекулярной организации клеток.
в) Комплекс Гольджи. Этот комплекс, называемый еще пластинчатым, был открыт итальянцем Камилло Гольджи еще в 1898 г. Он присутствует во всех клетках, кроме эритроцитов и сперматозоидов, и представляет собой систему дискообразных однослойных мембран (мембранных пузырьков или цистерн), локализующихся рядом с гладким эндоплазмати-ческим ретикулом и ядром (рис. 49). Часто в клетках обнаруживают несколько таких комплексов (диктиосом), размеры которых составляют 30-60 нм. Структурными молекулами, поддерживающими структуру цистерн, являются ферменты, вовлеченные в процессинг оли-госахаридов, белки, являющиеся аутоантителами, а также белки, являющиеся компонентами цитоскелета. Основная функция комплекса Гольджи заключается в том, что он является местом упаковки (уплотнения) белков, поступающих с рибосом, а также присоединения к белкам углеводов (образования гликопротеидов), а к полисахаридам — сульфатных групп с последующим транспортом их к другим клеточным структурам или за пределы клетки (экзоцитоз). В клетках печени этот комплекс участвует в выделении в кровь липопротеидов. Как отмечено выше, он участвует также и в формировании лизосом.
Цитоплазматический матрикс. Этот структурный компонент является основным веществом (цитоплазмой, гиалоплазмой) клетки. Первые электронномикроскопические изображения цитоплазмы были получены шведским ученым Ф. Шестрандом еще в 1955 г. Различают эктоплазму — вещество, располагающееся ближе к цитоплазматической мембране (твердое тело) и эндоплазму, отстоящую к центру клетки от эктоплазмы (более жидкое состояние) и представляющую собой цитозоль. Консистенция цитозоля приближается к гелю, В нем растворены многие ферменты и белки, обеспечивающие связывание и транспорт питательных веществ, микроэлементов и кислорода. Здесь же находятся аминокислоты и нуклеотиды, а также различные метаболиты (промежуточные продукты биосинтеза и распада макромолекул). Наконец, в цитозоле присутствуют различные коферменты, а также АТФ, АДФ, ионы ряда неорганических солей (K+, Mg2+, Са2+, C1-, НСО-2 3, НРО4-2), тРНК.
В цитоплазме содержатся микрофиламенты (нити) толщиной 4-5 нм и микротрубочки, представляющие собой полые цилиндрические структуры диаметром 25 нм, а также филаменты промежуточных размеров. Эти структуры составляют жесткую конструкцию (каркас) в клетке, называемую цитоскелетом и определяющую внешний вид и форму клеток. Микрофиламенты состоят из белка, сходного с сократительным белком актином.
Объединяясь, микрофиламенты формируют пучки, в которые входят дополнительные белки (анкерин, спектрин и другие). Основная функция микрофиламентов заключается в обеспечении сократительных процессов клеток, в упрочении мембран. Микротрубочки построены из белков a- и b-тубулина, а также g-тубулина. Для микротрубочек характерен ряд функций. Формирование микротрубочек происходит в интерфазе клеток в так называемых центрах организации микротрубочек (ЦОМ), которые «окружают» центриоли, в результате чего предполагают, что центриоли являются ЦОМ (рис. 50). В каждом ЦСУ содержится по 10-13 молекул g-тубулина и примерно по 7 молекул других белков, включая ди-мер a/b-тубулина. Эти белки формируют структуру, которая образует микротрубочный «ансамбль». Их значение до конца не выяснено, но предположительно заключается в том, что они обеспечивают перемещение клеточных органелл, включая хромосомы, внутри клеток.
В составе цитоплазмы обнаруживают ферменты, полисахариды, АТФ, тРНК, ионы Са, Na, К и других химических элементов.
Основная функция цитоплазматиче-ского матрикса заключается в том, что он является внутренней средой клетки, поддерживающей мембранные системы, орга-неллы и включения. В нем осуществляются гликолиз, активация аминокислот и другие реакции. Цитоскелет выполняет опорную функцию. Микрофиламенты способствуют упрочению мембранной системы, а микротрубочки, как отмечено выше, обеспечивают перемещение клеточных органелл и транспорт химических соединений из одних отсеков клетки в другие. Цитоскелет имеет значение также в делении клетки.
Клеточные органеллы. Эти структуры представлены ядром, хромосомами, ядрышком, центриолями, митохондриями, рибо-сомами, лизосомами. Они характерны за некоторыми исключениями как для клеток животных, так и для клеток растений.
а) Ядро в клетке имеет форму и размеры, зависящие от формы, размеров и функций клетки. В клетках эукариотических организмов содержится, как правило, по одному ядру, реже по два и более. В зрелом состоянии эритроциты млекопитающих и клетки ситовидных трубочек покрытосеменных растений лишены ядер, тогда как клетки скелетных мышц позвоночных и млечных сосудов растений являются многоядерными. Для инфузорий характерно наличие двух ядер — одно небольшое (микронуклеус) и одно крупное политенное (макронуклеус).
Обычно ядро имеет округлую, палочковидную, четковидную вытянутую и другие формы. Размеры его колеблются от 2 до 100 мкм, а объем составляет около 65 мкм3. Особенно крупные ядра характерны для половых клеток (размером до 500 мкм). Отношение объема ядра к объему цитоплазмы называют ядерно-плазменным отношением, которое у клеток всех типов обычно постоянно.
Строение ядра характеризуется чрезвычайной сложностью, хотя принципиально одинаково в клетках всех эукариотических организмов. В случае животных клеток ядро располагается в центре клетки. Напротив, в клетках растений оно имеет пристеночную локализацию. Однако положение, форма и размеры ядра могут меняться в зависимости от интенсивности метаболизма. В ядре содержатся хромосомы и ядрышки (см. ниже). Благодаря им ядро плотно заполнено ДНК, РНК и белками. Содержимое ядра иногда называют нуклеоплазмой или кариоплазмой. Она отделена от цитоплазмы ядерной мембраной, построенной из двух слоев (наружного и внутреннего) толщиной по 7 нм каждый и имеющей поры диаметром 40—100 нм. Поры занимают около 5% площади ядра. Через ядерные поры синтезируемая в ядре РНК выходит в цитоплазму, где она участвует в трансляции генетической информации (синтезе белков).
Ядерная мембрана и ядерные поры объединены с мембранной системой клетки, в результате чего клетка, по существу, канализирована различными проходами, обеспечивающими двухстороннее движение веществ в клетке, начиная от плазматической мембраны. В порах обнаружены глобулярные и фибриллярные белковые структуры. Количество пор зависит от вида клеток и обычно увеличивается в 8-периоде. Между мембранными слоями имеется так называемое перинуклеарное пространство шириной 20—60 нм. Наружная ядерная мембрана часто переходит в эндоплазматическую сеть. Когда клетки делятся, ядерная оболочка исчезает, а после деления восстанавливается. Ядерное вещество (кариоплазма) — плотный коллоид.
Важнейшая функция ядра заключается в том, что оно является центром управления в клетке, в нем происходит синтез ДНК, РНК и ядерных белков.
б) Хромосомы. Хромосомы располагаются в ядре. Их название введено в литературу В. Вальдейлом в 1883 г. Они имеют форму палочек, нитей, петель. Для каждой хромосомы характерны индивидуальные особенности, касающиеся длины и положения перетяжки (центромеры) по длине хромосомы. Каждая из митотичес-ких индивидуальных хромосом состоит из двух сестринских хроматид, удерживаемых центромерой. В зависимости от локализации центромер различают метацентрические, субметацентричес-кие, акроцентрические и телоцентрические хромосомы (рис. 51). Количество хромосом постоянно в ядрах соматических клеток, где они находятся в парах. Диплоидный набор хромосом называют кариотипом (от греч. caryon — ядро, type — форма, тип). Для разных организмов характерны разные по количеству диплоидные наборы хромосом. В зависимости от строения хромосомы человека классифицируют на группы.
Когда ядра соматических метафазных клеток окрашивают основными красителями, то при микроскопии препаратов в ядерной зоне обнаруживают глыбки и гранулы окрашенного вещества, получившего название хроматина. Именно из этого вещества во время деления клеток организуются хромосомы. При изучении таких микроскопических препаратов отмечают, что некоторые районы хромосом окрашены очень интенсивно, Другие слабее. Интенсивно окрашивающийся хроматин получил название гетерох-роматина, менее окрашивающийся — эухроматина. Последний содержит области, на которых очень активно синтезируется РНК.
Исследование хромосом типа ламповых щеток, содержащихся в овоцитах многих животных и политенных хромосомах, обнаруживаемых в клетках насекомых, показало, что они имеют петельную структуру. Эта структура характерна, вероятно, для организации хроматина у всех организмов.
В клетках многих видов организмов гетерохроматин концентрируется в основном в районах, фланкирующих центромеры.
Половые хромосомы часто почти полностью представлены гете-рохроматином. Гетерохроматином богаты районы повторов последовательностей ДНК (см. гл. X).
Когда гетерохроматин исследуют под электронным микроскопом, то отмечают, что он построен из плотноупакованных хрома-тиновых нитей диаметром 25 нм. Эухроматин составлен из менее плотноупакованных нитей, но такого же диаметра. Г
в) Ядрышко — это очень мелкая структура диаметром 1—5 мкм, которая локализуется в ядре. Количество ядрышек бывает разным — от одного до нескольких. Центральная часть в ядрышке представлена фибриллярной частью (цепи, ДНК- ядрышковые организаторы и рибонуклеопротеиды) и гранулярной (формирующиеся субъединицы рибосом). Ядрышко является местом синтеза рРНК. Матрицей для синтеза рРНК является ДНК ядрышкового организатора. Синтезируемая рРНК объединяется с белком и образующиеся рибонуклеопротеиды служат затем материалом для сборки из них субъединиц рибосом. Последние через ядерные поры направляются в цитоплазму, участвуя там в формировании рибосом, на которых происходит затем синтез белков.
г) Центриоли представляют собой гранулярные цилиндры диаметром около 0,15 мкм и длиной 0,5 мкм, локализующиеся парами в клетке около ядра. Эти пары называют диплосомами (центросома-ми). Структурно они представляют систему мембранных трубочек. Диплосома окружена другими микротрубочками, называемыми центросферой. Обе эти структуры образуют клеточный центр.
Функцией клеточного центра является организация цитоскеле-та клеток. Кроме того, центриоли участвуют в делении клеток, расходясь к полюсам клетки. Микротрубочки, протягивавшиеся взаимно, образуют митотическое веретено.
В клетках растений этой органел-лы нет.
д) Митохондрии присутствуют во всех клетках организмов, которые используют для дыхания кислород. В одной клетке может находиться 50-5000 митохондрий. Они имеют форму палочек, нитей или гранул (рис. 54), а их размеры достигают 7 мкм.
Митохондрии образованы двумя мембранами — наружной и внутренней, между которыми образуется пространство шириной 10—20 нм. Внутренняя мембрана формирует складки (кристы), которые погружены в матрикс, представляющий собой молекулярное содержимое митохондрий. Наружная мембрана проницаема для низкомолекулярных соединений. Проникновение веществ во внутреннее пространство (матрикс) митохондрий контролируется внутренней мембраной. Содержимое митохондрии составляют белки, фосфолипиды, ДНК, РНК и рибосомы, которых значительно меньше, чем рибосом, локализованных в цитоплазме. Наличие этих структур обеспечивает собственную митохондриальную белоксинтезирующую систему. Митохондрии способны к самовоспроизведению путем деления или почкования в период 8-фазы и других фаз клеточного цикла.
Митохондрии имеются также у простейших. В частности, ти-паносомы содержат по одной митохондрии.
Митохондрии представляют собой «силовые станции», в которых происходят основные процессы получения и накопления энергии путем окисления молекул пищи (окислительное фосфорилиро-вание) для образования АТФ. В матриксе содержатся ферменты, а реакции окисления проходят на поверхности внутренней мембраны. Энергетические потребности клеток определяют уровень размножения митохондрии.
В клетках растений вместо митохондрии содержатся пластиды. Среди пластид различают хлоропласты, которые содержат хлорофилл, лейкопласты (бесцветные пластиды), в которых происходит накопление крахмала, и хромопласты, в которых происходит синтез пигмента плодов. У пурпурных бактерий имеются хроматофоры.
Для хлоропластов характерно мембранное строение (рис. 55). Встречаясь в клетках в количестве около 40 экземпляров на клетку, они имеют своеобразную форму двояковыпуклых линз и размеры 5-10 мкм. Каждый хлоропласт окружен двойной мембраной. Наружная мембрана является гладкой, состоя из белковых субъединиц, а внутренняя — складчатой. Внутренняя мембрана содержит хлорофилл, а также ферменты, синтезирующие АТФ и органические соединения с помощью АТФ. Кроме того, в хлоро-пластах имеется некоторое количество ДНК и РНК небольших по размерам рибосом, что обеспечивает собственную хло-ропластную белоксинтезирующую систему.
Для пластид характерны переходы из одного типа в другой. Например, зеленые хлоропласты листьев переходят в хромопласты (осенью, когда изменяется окраска листьев), а лейкопласты зеленеющих клубней картофеля переходят в хлоропласты.
е) Рибосомы представляют собой гранулы, расположенные в ци-топлазматическом матриксе и связанные с мембранами цитоплаз-матической сети. Их размеры составляют 15—Збнм в диаметре. Они построены на одну треть из белка и на две трети из РНК (двух субъединиц). Количество рибосом в клетках очень большое. Например, в одной бактериальной клетке (кишечной палочке) их содержится около 6000 экземпляров. Рибосомы образуют группы, называемые полирибосомами.
Функции рибосом заключаются в том, что на них осуществляется синтез белков.
ж) Лизосомы представляют собой мельчайшие пузырьковидные образования, окруженные однослойной мембраной и содержащие ферменты. Их размеры составляют 0,2-0,8 мкм. Лизосомы обеспечивают изоляцию гидролитических ферментов клетки.
В лизосомах обнаружено более 40 различных гидролитических ферментов (протеиназ, нуклеаз, липаз, гликоидаз и др.), осуществляющих внутриклеточное расщепление макромолекул, проникающих в клетки посредством фагоцитоза и пиноцитоза. Лизосомы чрезвычайно разнообразны по строению и частным функциям. Предполагают, что они образованы мембранами комплекса Гольджи и представляют собой систему удаления из клеток конечных продуктов обмена. Повреждение лизосом сопровождается растворением клеток. Следовательно, благодаря лизосомам происходит защита клеток от собственных ферментов. В эволюционном плане они являются аналогами пищеварительных вакуолей одноклеточных организмов.
з) Пероксисомы (иногда их называют микротельцами) представляют собой образованные цистернами эндоплазматической сети пузырьки размером 0,3-1,5 мкм, ограниченные однослойной мембраной. Эти пузырьки содержат каталазу и некоторые оксидазы. Обильными по содержанию пероксисом являются эпителии печени и почек. Функция пероксисом заключается в том, что в них происходит разрушение клеточных перекисей и холестерина. Кроме того они содержатся также в клетках зеленых листьев растений. Известны также глиоксисомы, содержащиеся в проростках семян масличных растений.
Включения. В клетках животных включения представлены жировыми каплями, гранулами гликогена, зимогеновыми гранулами (депо ферментов в клетках поджелудочной железы). Особенно богаты жиром у млекопитающих клетки соединительной ткани. Гликогена много в клетках поперечно-полосатых мышц, печени и в нейронах. Встречаются также белковые включения в цитоплазме яйцеклеток, печени, в теле простейших. Следует отметить, что некоторые клетки животных содержат в качестве включений пигменты. Например, в клетках коркового вещества надпочечников накапливаются липохромы. Пигментами являются также гемоглобин и меланин, а также ретинин (в зрительном пурпуре сетчатки глаза).
В клетках растений, например, таких как картофель, злаковые, наиболее частым включением является крахмал. В качестве включений в клетках растений встречаются также жиры, содержание которых очень большое в семенах масличных культур. Очень широко распространены пигменты.
Участки (компартменты) цитоплазматического матрикса, лишенные мембран и клеточных органелл, получили название цитозолл.
4)Свойства клеточной мембраны
Кожа человека - это барьер, защищающий клетки, её мы и называем – клеточной мембраной. Она не позволяет компонентам клетки (цитоплазме) вытечь наружу. Главная задача клеточной мембраны - это удерживать клетку в целостности, при этом определять, что может попасть внутрь клетки, а что может оттуда выйти. Клетки любого организма имеют клеточные мембраны, даже клетки бактерий.
Строение клеточной мембраны
Состоит
клеточная мембрана из бинарного
ряда липидов.
Располагаются молекулы липидов в два
ряда и каждый ряд точно такой же, как
предыдущий. Структуру молекулы липида
- эти две части единого целого, как раз
и отображают. Ещё эти две части единого
целого называют – гидрофобной
(водонепроницаемой) и гидрофильной
секциями.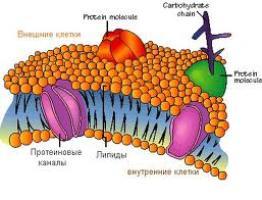
Гидрофобная секция не любит воду и подобных воде молекул, благодаря бинарному слою липидов выступает вроде защитного механизма.
Гидрофильная секция напротив способна притягивать воду и подобные воде молекулы, после чего выталкивает их наружу. В итоге получается такая базовая жидкая мозаичная модель.
Жидкостно-мозаичная модель
Открыли жидкую мозаичную модель клеточной мембраны в 1972 году. Эта модель демонстрирует структуру размещения протеинов внутри или на биполярном слое липидов. Размещаются протеины в хаотичном порядке, при этом получается мозаика протеинов. Протеины пересекающие бинарный ряд липидов, играют важную роль в транспортировке маленьких молекул через мембрану.
-
Нашли ошибку в тексте? Выделите ее и еще
несколько слов, нажмите Ctrl + Enter
-
Вам не понравилась статья или качество
поданной информации? - напишите нам!![]()
Функции клеточной мембраны
В
результате того, что клеточная мембрана
имеет среду полупроницаемую, то только
некоторые виды самых мелких молекул
способны проникнуть внутрь и наружу
клетки сквозь мембрану. Называется
данный процесс – диффузией. Но для того,
чтобы такое действие произошло, мембрана
должна быть вроде открытых дверей, то
есть проницаемой, чтобы маленькая
молекула смогла проникнуть сквозь неё.
В том случае, когда мембрана непроницаема,
маленькая молекула не имеет возможности
проникнуть, здесь можно сказать «двери
закрыты». Однако следует учесть, что
молекула может переместиться только
из переполненной области, в более
свободную. Например, аминокислота желает
пробраться через мембрану к самой
клетке, при этом клеточная мембрана
открыта для неё, это может произойти в
том случае если концентрация аминокислоты
за пределами клетки больше, чем в самой
клетки. Только при таких условиях
произойдёт процесс диффузии.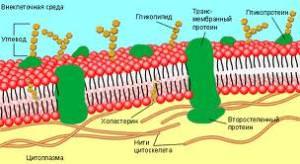
Следующий процесс, в котором главную роль играет клеточная мембрана - это так называемая помощь при транспортировке. Некоторые маленькие молекулы в мембране которых разместились протеины, способны помочь при пересечении мембраны. Вроде протеинового эскорта сквозь неё. Данный процесс напоминает процесс диффузии, так как протеиновая помощь при пересечении мембраны происходит, когда молекулы переходят из области высокой концентрации, с целью понизить её. Или же просто из области высокой концентрации молекул в область менее загруженную.
Опять же, молекулы способны пересекать клеточную мембрану способом активной транспортировки. Такой вид транспортировки представляет собой переход молекул из области с низкой концентрации в противоположную область с высокой концентрацией. Такой процесс транспортировки, противоположный диффузии и посреднической транспортировки, осуществляется против правил концентрационной направленности. Для того, чтобы такие передвижения осуществлять необходима концентрация энергии в форме аденозинтрифосфата (АТФ). АТФ- это нуклеотид, который состоит из аденина, рибозы и трёх остатков фосфорной кислоты. Он является универсальным накопителем и переносчиком химической энергии, известным для всех организмов и клеток. АТФ применяется для некоторых клеточных действий, начиная движениями и заканчивая репродукцией.
5)БИОЛОГИЧЕСКИЕ МЕМБРАНЫ
Все живые клетки отделены от окружающей среды поверхностью называемой клеточной мембраной. Кроме того, для эукариотов характерно образование внутри клеток нескольких компартментов. Они представлены рядом субклеточных органелл, ограниченных мембранами, например, ядро и митохондрии. Мембраны представляют собой не только статически организованные поверхности раздела, но и включают активные биохимические системы, отвечающие за такие процессы, как избирательный транспорт веществ внутрь и наружу клетки, связывание гормонов и других регуляторных молекул, протекание ферментативных реакций, передача импульсов нервной системы и т.д. Существуют различные типы мембран, отличающиеся по выполняемым функциям. Функции мембран обусловлены их строением.

Функции мембран
Химический состав
Мембраны состоят из липидных и белковых молекул, относительное количество которых варьирует (от 1/5 - белок + 4/5 - липиды до 3/4 - белок + 1/4 – липиды) у разных мембран. Углеводы содержатся в форме гликопротеинов, гликолипидов и составляют 0,5-10% вещества мембраны.
Липиды мембран
Основная часть липидов в мембранах представлена фосфолипидами, гликолипидами и холестерином. Строение этих липидов представлено на рисунке:
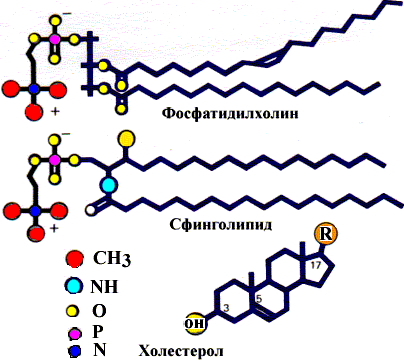
Строение липидов мембран
Липиды мембран имеют в структуре две различные части: неполярный гидрофобный «хвост» и полярную гидрофильную «голову». Такую двойственную природу соединений называют амфифильной. Липиды мембран образуют двухслойную структуру. Каждый слой состоит из сложных липидов, расположенных таким образом, что неполярные гидрофобные «хвосты» молекул находятся в тесном контакте друг с другом. Так же контактируют гидрофильные части молекул. Все взаимодействия имеют нековалентный характер. Два монослоя ориентируются «хвост к хвосту» так, что образующаяся структура двойного слоя имеет внутреннюю неполярную часть и две полярные поверхности. Белки мембран включены в липидный двойной слой двумя способами:
связаны с гидрофильной поверхностью липидного бислоя - поверхностные мембранные белки
погружены в гидрофобную область бислоя - интегральные мембранные белки.
Поверхностные белки своими гидрофильными радикалами аминокислот связаны нековалентными связями с гидрофильными группами липидного бислоя. Интегральные белки различаются по степени погруженности в гидрофобную часть бислоя. Они могут располагаться по обеим сторонам мембраны и либо частично погружаются в мембрану, либо прошивают мембрану насквозь. Погруженная часть интегральных белков содержит большое количество аминокислот с гидрофобными радикалами, которые обеспечивают гидрофобное взаимодействие с липидами мембран. Гидрофобные взаимодействия поддерживают определенную ориентацию белков в мембране. Гидрофильная выступающая часть белка не может переместиться в гидрофобный слой. Часть мембранных белков ковалентно связаны с моносахаридными остатками или олигосахаридными цепями и представляют собой гликопротеины. Примеры расположения белков и липидов в мембране представлены на рисунке:
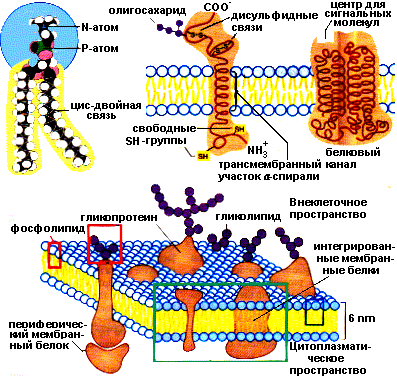
Структура плазматической мембраны
Асимметрия мембран
Хотя каждый монослой образован из липидов, ориентированных одинаковым образом, тем не менее, липидный состав монослоев различен. Например, в плазматической мембране эритроцитов фосфатидилхолины преобладают в наружном слое, а фосфатидилсерины во внутреннем слое мембраны. Углеводные части белков и липидов располагаются на наружной части мембраны. Кроме того, поверхности мембраны отличаются по составу белков. Степень такой асимметрии мембран различна у разных типов мембран и может меняться в процессе жизнедеятельности клетки и ее старения. Подвижность (жесткость) и текучесть мембран также зависят от ее состава. Повышенная жесткость обуславливается увеличением соотношения насыщенных и ненасыщенных жирных кислот, а также холестерина. Физические свойства мембран зависят от расположения белков в липидном слое. Липиды мембран способны к диффузии в пределах слоя параллельно поверхности мембраны (латеральная диффузия). Белки тоже способны к латеральной диффузии. Поперечная диффузия в мембранах сильно ограничена.