

Молекулярная биология клетки Глава 4
.pdfГБОУ ВПО «КАЗАНСКИЙ ГОСУДАРСТВЕННЫЙ МЕДИЦИНСКИЙУНИВЕРСИТЕТ» МИНИСТЕРСТВАЗДРАВООХРАНЕНИЯ РФ
КАФЕДРА МЕДИЦИНСКОЙ БИОЛОГИИ И ГЕНЕТИКИ
МОЛЕКУЛЯРНАЯ БИОЛОГИЯ КЛЕТКИ
Часть 4. Половое размножение. Гаметы. Мейоз
Учебное пособие
Казань 2015
УДК 611:018(075.8) ББК 28.05+28.06
Печатается по решению Центрального координационно-методического совета
Казанского государственного медицинского университета.
Авторы:
Исламов Р.Р., Волков Е.М., Кошпаева Е.С., Пахалина И.А., Колочкова Е.В., Бойчук Н.В.
Под редакцией д.м.н. профессора Исламова Р.Р.
Рецензенты:
Заведующий кафедрой гистологии, цитологии и эмбриологии КГМУ проф. Челышев Ю.А.
Заведующая кафедрой гистологии, цитологии и эмбриологии РНИМУ им. Н.И. Пирогова проф. Глинкина В.В.
Молекулярная биология клетки.Часть 4. Половое размножение. Гаметы. Мейоз. Учебное пособие, переработанное и дополненное / Исламов Р.Р., Волков Е.М., Кошпаева Е.С., Пахалина И.А., Колочкова Е.В., Бойчук Н.В. – Казань: КГМУ, 2015. – 37 с.
Учебное пособие составлено в соответствии с Государственным образовательным стандартом высшего профессионального образования (ФГОС) и типовой Учебной программой по дисциплине «Биология». В 4 части Учебного пособия «Молекулярная биология клетки» «Половое размножение. Гаметы. Мейоз» изложены цели и задачи данной темы, указаны формируемые компетенции, приведён теоретический обзор изучаемого материала, изложенный в лаконичной, рубрицированной форме и имеющий медицинскую направленность. Практические навыки включают изучение микропрепаратов мужских и женских половых клеток млекопитающих, микропрепаратов мейоза яйцеклетки конской аскариды, а также протокол лабораторной работы по двигательной активности живых сперматозоидов лягушки. Пособие содержит типовые тестовые вопросы, вопросы самоконтроля, справочник терминов.
Учебное пособие предназначено для студентов младших курсов медицинских факультетов и медицинских вузов.
© Казанский государственный медицинский университет, 2015
Часть 4. Половое размножение.
Гаметы. Мейоз
Целью публикации настоящего учебного пособия является систематизация знаний о размножении живых организмов. В учебное пособие включена информация о принципах получения эмбриональных стволовых клеток из гамет и клонирования.
Задачей учебного пособия является ознакомление студентов со строением половых клеток и основными этапами гаметогенеза на примере млекопитающих. Особое внимание уделяется усвоению биологической роли мейоза и молекулярных механизмах рекомбинативной изменчивости.
Формируемые компетенции:
•Лечебный факультет, дисциплина «Биология» — ОК1, ПК31
•Педиатрический факультет, дисциплина «Биология» — ОК1, ПК31
•Стоматологический факультет, дисциплина «Биология» — ОК1, ПК26
•Медико-профилактический факультет, дисциплина «Биология, экология» — ОК7, ПК6
•Фармацевтический факультет, дисциплина «Биология» — ОК1, ПК48
•Медико-биологический факультет, дисциплина «Биология» — ОК1, ПК27
Студент должен знать:
1.Строение яйцеклетки млекопитающих и стадии овогенеза.
2.Строение сперматозоида млекопитающих и стадии сперматогенеза.
3.Механизм рекомбинативной изменчивости.
4.Принципы клонирования.
Студент должен уметь:
1.Зарисовывать изучаемые микропрепараты мужских и женских половых клеток.
2.Идентифицировать различные стадии мейоза по микропрепаратам, микрофотографиям, рисункам и схемам.
4
3. Оценивать двигательную активность сперматозоидов.
Студент должен владеть:
1.Медико-функциональным понятийным аппаратом и терминологией по данной теме.
2.Навыками приготовления временного препарата живых сперматозоидов.
3.Навыками описания хромосомных нарушений при мейозе.
Оснащение занятия:
1.Мультимедийная презентация по теме.
2.Таблицы:
•типы яйцеклеток
•строение сперматозоида
•схема мейоза
3.Микроскопы:
•прямой световой монокулярный
•прямой световой бинокулярный
4.Микропрепараты:
•яйцеклеткакошки
•сперматозоиды морской свинки
•стадии мейоза яйцеклетки конской аскариды
5.Студенческий лабораторный набор для приготовления временного препарата живых сперматозоидов лягушки.
Хронологическая карта занятия:
1.Организационная часть.
2.Тестовый контроль базового уровня знаний.
3.Объяснение практического задания.
4.Самостоятельная работа.
5.Проверка выполненных работ в альбомах.
6.Контроль конечного уровня знаний.
7.Установка задания для подготовки к следующей теме.
5
ТЕОРЕТИЧЕСКИЙОБЗОР Размножение — свойство всех живых организмов, обеспечиваю-
щее воспроизведение себе подобных форм жизни. Различают бесполое и половое размножение. В основе обеих форм размножения лежит единый механизм деления материнской клетки на две дочерние. Исключением является репликация вирусных частиц, характеризующаяся многократным копированием наследственного материала в инфицирован-
ной клетке.
Бесполое размножение — эволюционно более древнее и простое воспроизводство по сравнению с половым. У эукариот бесполое размножение происходит за счёт митотического деления соматических клеток без участия гамет (половых клеток). Поэтому образующееся потомство генетически идентично родительской особи. Бесполое размножения встречается как среди одноклеточных (сине-зелёные водоросли, простейшие), таки многоклеточных (грибы, растения, кольчатые черви, гидры, морские звёзды) эукариот, но происходит разными способами: вегетативным размножением (деревья, кустарники), почкованием (гидры, кораллы), спорообразованием (грибы, мхи, папоротники) или делением целого организма на части (плоские, кольчатые черви), из которых
образуются целостные организмы, сходные с родительской особью. Половое размножение характерно для диплоидных организмов, у
которых соматические клетки содержат двойной набор хромосом (по одному от каждого родителя) и связано с образованием мужских (сперматозоидов) и женских (яйцеклеток) половых клеток с гаплоидным набором хромосом. При слиянии двух половых гаплоидных клеток восстанавливается диплоидный набор хромосом, а в результате смешивания геномов двух индивидуальных организмов появляются потомки, генетически отличающиеся, как друг от друга, так и от своих родителей.
Конкуренция за полового партнёра между особями одного пола влечёт за собой выборочное спаривание (половой отбор). По существу, гене-
тическая рекомбинация в ходе созревания половых клеток (мейоза) и половой отбор являются элементами естественного отбора в эволюции вида.
ПОЛОВЫЕ КЛЕТКИ
Сперматозоид
Открытие сперматозоидов принадлежит голландскому натуралисту Антони ван Левенгуку. В 1679 г. Левенгук доложил о «семенных зверьках» в семенной жидкости человека Лондонскому королевскому обще-
6
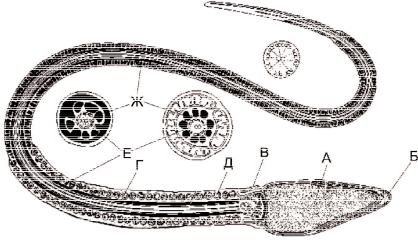
ству. Термин «сперматозоид» ввёл академик Петербургской Академии Карл Эрнст фон Бэр в начале XIX века. В сперматозоиде человека различают головку, тело и хвостик/жгутик (flagellum) (рис. 4-1). Яйцевидная, сплющенная с двух сторон головка содержит ядро с гаплоидным набором хромосом и акросому с ферментами для растворения прозрачной оболочки яйцеклетки (гиалуронидаза, протеазы, гликозидазы, липазы, нейраминидаза, фосфатазы). Тело сперматозоида имеет утолщение за счёт спирально закрученной нитевидной митохондрии. Между телом и головкой расположены дистальная (центр организации аксонемы) и проксимальная, локализующаяся рядом с ядром, центриоли. В осевой части тела и хвостика, начиная от дистальной центриоли, находится сократительная органелла — аксонема, образованная микротрубочками по принципу 9+2 (девять пар микротрубочек по периферии и одна пара в центре) (рис. 4-2). Снаружи от аксонемы расположено 9 фибрилл, состоящих из электроноплотного материала. Фибриллы начинаются от шейки, проходят через тело и заканчиваются на уровне се-
Рис. 4-1. Сперматозоид человека. Головка содержит ядро (А) и акросому(Б). В шейке (В) присутствуют дистальная и проксимальная центриоли. Промежуточный отдел (Г) содержит спирально закрученную нитевидную митохондрию (Д). В осевой части промежуточного отдела и почти вдоль всего хвоста расположена аксонема (Е). В хвосте снаружи от аксонемы находятся наружные плотные фибриллы и фибриллярный футляр (Ж) [из Fawcett D.W., 1975].
7
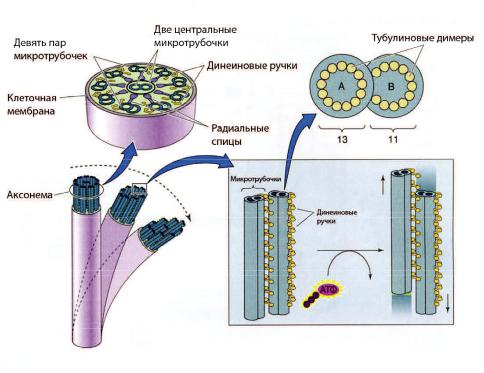
Рис. 4-2. Тубулин-динеиновый хемомеханический преобразователь. Глобулярная головка динеина обладает АТФазной активностью. При расщеплении АТФ она скользит по поверхности микротрубочки (субфибрилла B) соседней пары по направлению к её (–)-концу. Этот механизм аналогичен скольжению элементов актомиозинового хемомеханического преобразователя в мышце [из Boron W.F., Boulpaep E.L., 2003].
редины хвостика. Фибриллы в пределах хвостика окружены фибриллярным футляром из электроноплотного материала, не доходящего до концевого отдела хвостика.
Вэякуляте здорового мужчины содержится примерно от 150 до 600 млн сперматозоидов, 100 тыс. из них попадут в полость матки и примерно 200 достигнут воронки маточной трубы, где происходит встреча яйцеклетки и сперматозоида. В женских половых путях сперматозоиды сохраняют способность к оплодотворению максимально до 2 суток.
В23% случаев мужского бесплодия обнаруживается патозооспермия (отсутствие или недостаточное количество сперматозоидов, морфологические дефекты, снижение подвижности сперматозоидов).
8
Ядро
Ядро сперматозоида содержит гаплоидный набор хромосом (22+Х или 22+Y). Слияние сперматозоида 22+Х с яйцеклеткой 22+Х даст начало женскому организму. Слияние сперматозоида 22+Y с яйцеклеткой 22+Х даст начало мужскому организму.
Y-хромосома представлена во всех ядрах клеток мужского организма. При мейозе Y-хромосома рекомбинирует с Х-хромосомой. Участок Y-хромосомы, гомологичный с Х-хромосомой, расположен на концах Y-хромосомы (псевдоаутосомный регион и теломера) и занимает около 5% её длины. Приблизительно 95% Y-хромосомы не имеют гомологичных участков и не рекомбинируют. В этих участках локализуются гены, функционирующие только в мужском организме.Y-хромосома содержит около 200 генов, среди которых 72 гена кодируют белки. Признаки, наследуемые с Y-хромосомой, называют голандрическихми.
Нерекомбинантные геныY-хромосомы, присутствующие на Х-хро- мосоме
AMELY/AMELX (амелогенин) RPS4Y1/RPS4Y2/RPS4X (рибосомный белок S4)
Нерекомбинантные гены Y-хромосомы, отсутствующие на Х-хро- мосоме
AZF1 (азооспермии фактор 1)
BPY2 (основной белокY-хромосомы)
DAZ1 (удаляется при азооспермии)
PRKY (Y-сцепленная протеин киназа) SRY (регион, определяющий пол) TSPY (специфичный для яичка белок) ZFY (белок цинковых пальцев)
Аксонема
Движение жгутика сперматозоида и ресничек мерцательных клеток обеспечивает тубулин-динеиновый хемомеханический преобразователь (тубулиновый молекулярный мотор) в составе аксонемы. Аксонема формируется путём самосборки. Матрицей для сборки служит центриоль или базальное тельце. Аксонема (рис. 4-2) состоит из 9 периферических пар микротрубочек и двух расположенных центрально одиночных микротрубочек. В каждой периферической паре различают субфибриллу А, содержащую 10–11 тубулиновых протофиламентов, и субфибриллу В, содержащую 13 протофиламентов. Смежные пары микротрубочек соединены междусобой эластичным белком нексином. С субфибриллой А связаны наружные и внутренние ручки. В их состав входит белок дине-
9
ин, содержащий глобулярные головки, соединённые с гибкой фибриллярной частью молекулы. Основание фибриллярной части вплетено в микротрубочку(субфибрилла A).
Дефект гена DNAI1 (аxonemaldynein intermediate chain 1) — причина отсутствия одной или обеих динеиновых ручек. Клинически дефект характеризуется рецидивирующими инфекциями дыхательных путей, бесплодием у мужчин, пониженной фертильностью у женщин; наследуется по аутосомно-рецессивному типу. Более половины больных с подобным синдромом имеет situs viscerus inversus — транспозицию внутренних органов (сердце справа, печень слева и т.д.), что в совокупности описывает синдром Картагенера.
Рецепторный аппарат
Рецептор β1,4-галактозилтрансферазаI(GalT I)в мембране головки сперматозоида. GalT I взаимодействует с олигосахаридами гликопроте-
ина ZP3 прозрачной оболочки.
Обонятельные рецепторы hOR17-4, локализованные в мембране хвоста сперматозоида, отвечают за хемотаксис. Связанные с G- белком (Gaolf) рецепторы hOR17-4 активируют аденилатциклазу (mAC III), запускающую внутриклеточный каскад на повышение уровня ионов кальция в цитоплазме хвоста. Повышение концентрации кальция модулирует движение хвоста сперматозоида, изменяя его направление. Пахучее вещество bourgeonal, имеющее запах ландыша, специфически связывается с рецептором hOR17-4 и является сильным хемоатрактантом для сперматозоидов. На основе химической формулы бурженаль ведутся поиски естественных хемоатрактантов сперматозоидов.
ЯЙЦЕКЛЕТКА
Зрелая женская половая клетка имеет шарообразную форму диаметром 100-150 µм, не способна к активному движению. Центрально расположенное округлое ядро с выраженным ядрышком содержит гаплоидный набор хромосом (22+Х). В цитоплазме яйцеклетки, кроме типичных органелл, содержится небольшое количество желточных включений (lecythos) с питательными веществами (белки, жиры, углеводы, минеральные соли), используемыми на начальных этапах развития зародыша, а под плазмолеммой локализуются кортикальные гранулы, содержащие ферменты (протеазы). Дегрануляция (кортикальная реакция) происходит в ответ на проникновение сперматозоида в яйцеклетку. Протеазы модифицируют прозрачную оболочку, которая превращается в обо-

10
лочку оплодотворения и препятствует смертельной для яйцеклетки полиспермии.
Прозрачная оболочка
По мере развития яйцеклетки в ней происходят синтез и последующая секреция гликопротеинов, постепенно формирующих zonapellucida (рис. 4-3). Зрелая прозрачная оболочка содержит густую сеть тонких нитей, состоящую из гликопротеинов (главным образом, белки ZP); один из них (ZP3) — главный гликопротеин для связывания рецептора b1,4- галактозилтрансфераза I (GalT I) в мембране головки сперматозоида. Инактивация или отщепление О-олигосахарида от молекулы ZP3 блокирует связывание сперматозоида с яйцеклеткой (образование оболочки оплодотворения). ZP2 — вторичный рецептор сперматозоида, обеспечивающий дополнительное связывание гамет. Связывание сперматозоида с гликопротеинами ZP является сигналом к акросомной реакции.
Рис. 4-3. Оболочки яйцеклетки млекопитающего. Снаружи от плазмолеммы овоцита располагается прозрачная оболочка, содержащая гликопротеины ZP1, ZP2 и ZP3. Лучистый венец образуют 2–3 слоя фолликулярных клеток снаружи от прозрачной оболочки. Фолликулярные клетки между собой и с овоцитом связаны щелевыми контактами для осуществления межклеточных коммуникаций. Щелевые контакты найдены на концах отростков клеток лучистого венца, которые проникают через прозрачную оболочку и вступают в контакт с поверхностью овоцита. [из Gilbert S.F., 2000 после Wassarman P.M., 1989 и по Baker T.G, 1972 из Ham A.G., Veomett M.J., 1980].