

ответы к экзамену по гистологии
.docx22. Строение нервов Нерв - покрытая оболочкой структура, состоящая из пучка нервных волокон (главным образом, представленных аксонами нейронов) и поддерживающей их нейроглии. Периферический нерв состоит из нескольких пучков аксонов, покрытых оболочками из Шванновских клеток, а также несколькими соединительно-тканными оболочками: эндоневрий покрывает каждый миелинизированный аксон, несколько таких аксонов объединяются в пучки, покрытые периневрием. Несколько пучков, вместе с кровеносными сосудами и жировыми включениями, покрыты общей оболочкой, эпиневрием, и составляют нерв. Нервы подразделяются на: 1) чувствительные (аферентные) - состоят из дендритов чувствительных нейронов, проводят импульс из рецепторов в ЦНС. 2) смешанные - состоят из дендритов и аксонов, проводят импульс в двух направлениях (из рецептора в ЦНС и наоборот) 3) движения (эферентные) - состоят из аксонов нейронов движения, проводят импульс из ЦНС в исполнительные органы (мускулы и железы) Нервы образованы многочисленными пучками мякотных и безмякотных нервных волокон, которые объединяются в нервные стволы и изолируются соединительной тканью. Мякотные (миелиновые) волокна имеют оболочку из жироподобного вещества - миелина. Эту оболочку образуют шванновские глиальные клетки, закручивающиеся вокруг аксона. Между Шванновскими клетками - свободные участки аксона. Их называют перехватами Ранвье. Безмякотные (миелиновые) волокна не имеют мякотной оболочки, но все равно защищены шванновскими клетками. В цитоплазму этих клеток как бы вставлено несколько волокон.
23. Строение и роль в клетке фибриллярных структур Фибриллы - нитевидные структуры в клетках и тканях животных и растительных организмов. Состоят из протофибрилл, белка актина. В мышечных клетках содержат также миозин. К фибриллам относят: различные по происхождению и функциям, значению образования, видимые в световом микроскопе коллагеновые и эластичные волокна, миофибриллы, нейрофибриллы, тонофибриллы и обнаруживаемые электронномикроскопически протофибриллы мышц, цитоплазматические микрофибриллы. Фибриллы выполняют в клетке двигательную или скелетную функцию.
24. Строение серого и белого вещества спинного мозга На поперечном разрезе в спинном мозге выделяют внутреннее серое вещество (тела нервных клеток и их отростки), окружающее спинномозговой канал, и внешнее белое вещество (миелинизированные нервные волокна). На протяжении спинного мозга меняется отношение серого и белого вещества. Серое вещество представлено наименьшим количеством клеток в грудном отделе. Наибольшим - в поясничном. Серое вещество: На срезе спинного мозга большинства позвоночных серое вещество напоминает по форме бабочку (в зарубежной литературе обычно такое сравнение не употребляется). В нем выделяют передние и задние рога, в грудных отделах (у человека от 8 шейного до 2 поясничного) выражены боковые рога. В объеме серое вещество образует так называемые столбы. Серое вещество спинного мозга содержит различные нервные элементы, эти элементы могут располагаться диффузно, а могут быть собранными в виде ядер. Обычно выделяют такие ядра, как: 1) желатинозная субстанция, или вещество Роланда - находится в заднем роге, образовано мелкими нейронами, отвечает за проведение болевой и температурной информации; 2) собственное ядро заднего рога, или промежуточное ядро Кахаля - находится ниже желатинозной субстанции, состоит из вставочных нейронов, участвует в рефлексах спинного мозга, также отвечает за межсегментарные связи, не имеет трактов, выходящих за пределы спинного мозга; 3) ядро Кларка - расположено рядом с центральным каналом в основании заднего рога, сомы нейронов крупные, округлые, отправляет информацию от рецепторов мышц и сухожилий в мозжечок по двум спиномозжечковым трактам (Флешига и Говерса); 4) боковое ядро - расположено в боковых рогах сегментов с c 8 (восьмой шейный) до l 2 (3) (второй или третий поясничный), несет GVE-компонент (иннервирует вегетативные ганглии). Аналогичное образование есть в s2-s4 сегментах (со второго по четвертый крестцовые), но не в боковых рогах за их отсутствием; 5) моторные ядра - расположены в переднем роге, среди крупных альфа-мотонейронов лежат мелкие гамма-мотонейроны. В 1952 году шведский анатом Брор Рексед предложил разделять серое вещество на десять пластин (слоев), различающихся по структуре и функциональной значимости составляющих их элементов. Эта классификация получила широкое признание и распространение в научном мире. Пластины принято обозначать римскими цифрами.
Пластины с I по IV образуют головку дорсального рога, которая является первичной сенсорной областью. I пластина образована многими мелкими нейронами и крупными веретеновидными клетками, лежащими параллельно самой пластине. В нее входят афференты от болевых рецепторов, а также аксоны нейронов II пластины. Выходящие отростки контрлатерально (то есть, перекрестно - отростки правого заднего рога по левым канатикам и наоборот) несут информацию о болевой и температурной чувствительности в головной мозг по передним и боковым канатикам (спиноталамический тракт). II и III пластины образованы клетками, перпендикулярными к краям пластин. Соответствуют желатинозной субстанции. Обе афферируются отростками спиноталамического тракта и передают информацию ниже. Участвуют в контроле проведения боли. II пластина также отдает отростки к I пластине. IV пластина соответствует собственному ядру. Получает информацию от II и III пластин, аксоны замыкают рефлекторные дуги спинного мозга на мотонейронах и участвуют в спиноталамическом тракте. V и VI пластины образуют шейку заднего рога. Получают афференты от мышц. VI пластина соответствует ядру Кларка. Получает афференты от мышц, сухожилий и связок, нисходящие тракты от головного мозга. Из пластины выходят два спиномозжечковых тракта: 1) тракт Флешига (вариант: Флексига) - выходит ипсилатерально (то есть в канатик своей стороны) в боковой канатик; 2) тракт Говерса - выходит контрлатерально в боковой канатик. VII занимает значительную часть переднего рога. Почти все нейроны этой пластины вставочные (за исключением эфферентных нейронов бокового ядра). Получает афферентацию от мышц и сухожилий, а также множество нисходящих трактов. Аксоны идут в IX пластину. VIII пластина расположена в вентро-медиальной части переднего рога, вокруг одной из частей IX пластины. Нейроны ее участвуют в проприоспинальных связях, то есть связывают между собой разные сегменты спинного мозга. Пластина IX не едина в пространстве, ее части лежат внутри VII и VIII пластин. Она соответствует моторным ядрам, то есть является первичной моторной областью, и содержит мотонейроны, расположенные соматотопически (то есть представляет собой «карту» тела), например, мотонейроны мышц-сгибателей залегают обычно выше мотонейронов мышц-разгибателей, нейроны, иннервирующие кисть - латеральнее, чем иннервирующие предплечье, и т. д. X пластина расположена вокруг спинального канала, и отвичает за комиссуральные (между левой и правой частями спинного мозга) и другие проприоспинальные связи. Белое вещество: Белое вещество окружает серое. Борозды спинного мозга разделяют его на канатики: передние, боковые и задние. Канатики представляют собой нервные тракты, связывающие спинной мозг с головным. Самой широкой и глубокой бороздой является передняя срединная щель, разделяющая белое вещество между передними рогами серого вещества. Напротив неё - задняя срединная борозда. По паре латеральных борозд идут соотвественно к задним и передним рогам серого вещества. Задний канатик разделяют пара промежуточных борозд, образуя два восходящих тракта: ближний к задней срединной борозде нежный, или тонкий пучок, и более латеральный клиновидный пучок. Внутренний пучок, тонкий, поднимается с самых нижних отделов спинного мозга, клиновидный же образуется только на уровне грудного отдела.
25.
Центриоли, реснички и жгутики. Их строение
и роль в клетке:
Центриоли
- постоянные структуры всех животных и
некоторых растительных клеток, основная
часть клеточного центра. Центриоли
(обычно их две) лежат вблизи ядра. Каждая
центриоль построена из цилиндрических
элементов (микротрубочек), образованных
в результате полимеризации белка
тубулина. Девять триплетов микротрубочек
расположены по окружности.
Функции
центриолей:
Центриоли
принимают участие в формировании
цитоплазматических микротрубочек во
время деления клетки и в регуляции
образования митотического веретена. В
клетках растений центриолей нет, и
митотическое веретено образуется там
иным способом. Кроме того, ученые
полагают, что ферменты клеточного центра
принимают участие в процессе перемещения
дочерних хромосом к разным полюсам в
анафазе митоза.
Реснички - органеллы,
представляющие собой тонкие (диаметром
0,1 - 0,6 мкм), короткие (5 - 15 мкм) структуры
на поверхности эукариотических клеток.
Могут быть как подвижны, так и нет: в
этом случае играют роль рецепторов.
Характерны для инфузорий, у некоторых
многоклеточных беспозвоночных (ресничные
черви, личинки кишечнополостных и губок)
ими покрыта вся поверхность тела, у
позвоночных и человека клетки с ресничками
расположенны лишь в некоторых местах:
на эпителии дыхательных путей, евстахиевых
трубах, семявыносящих канальцах,
яйцеводов, матки.
Ультраструктура
ресничек и жгутиков идентична. Снаружи
они покрыты трёхслойной мембраной,
переходящей в поверхностную мембрану
клетки. В центре помещаются тянущиеся
вдоль всей реснички 2 центральные
трубчатые фибриллы и 9 периферических,
каждая из которых двойная. В поверхностных
слоях цитоплазмы клетки каждая ресничка
берёт начало от кинетосомы (базального
тельца), имеющей сходную с ресничкой
структуру, но лишённую центральных
фибрилл. Периферические фибриллы
обусловливают движение реснички, тогда
как центральные играют, по-видимому,
опорную, а возможно, и проводящую
возбуждение роль.
Жгутик - поверхностная
структура, присутствующая у многих
прокариотических и эукариотических
клеток и служащая для их движения в
жидкой среде или по поверхности твёрдых
сред. Жгутики прокариот и эукариот резко
различаются: бактериальный жгутик имеет
толщину 10 - 20 нм и длину 3 - 15 мкм, он
пассивно вращается расположенным в
мембране мотором; жгутики же эукариот
толщиной до 200 нм и длиной до 200 мкм, они
могут самостоятельно изгибаться по
всей длине. У эукариот часто также
присутствуют реснички, идентичные по
своему строению жгутику, но более
короткие (до 10 мкм).
Жгутики
прокариот:
Жгутики
бактерий состоят из трёх субструктур:
1)
Филамент (фибрилла, пропеллер) - полая
белковая нить толщиной 10 - 20 нм и длиной
3 - 15 мкм, состоящая из флагеллина,
субъединицы которого уложены по спирали.
Полость внутри используется при синтезе
жгутика - он происходит в направлении
от ЦПМ. По полости к собираемому в
настоящий момент участку переносятся
субъединицы флагеллина.
2) Крюк - более
толстое чем филамент (20-45 нм) белковое
(не флагеллиновое) образование.
3)
Базальное тело (трансмембранный
мотор).
Жгутикование
бактерий:
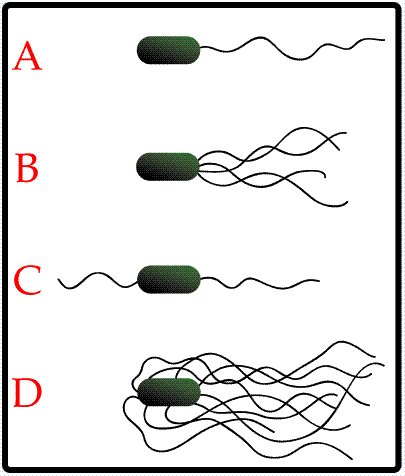 Описание
рисунка: A - монотрихиальное, B -
лофотрихиальное, C - амфитрихиальное, D
– перитрихиальное.
Описание
рисунка: A - монотрихиальное, B -
лофотрихиальное, C - амфитрихиальное, D
– перитрихиальное.
Базальное тело и механизм его работы: Базальное тело представляет собой систему колец, находящихся в ЦПМ и клеточной стенке бактерий. Два внутренних кольца - M и S-кольца (сейчас чаще рассматриваются как единое MS-кольцо) - являются обязательными элементами, причём M-кольцо находится в ЦПМ, а S - в периплазме грамотрицательных и пептидогликановом слое грамположительных бактерий. Ещё два кольца - P и L - есть только у грамотрицательных бактерий, они расположены в пептидогликановом слое и наружной мембране соответственно, неподвижны и лишь направляют стержень ротора мотора. Вокруг MS-кольца расположены статоры - белковые комплексы MotA4/MotB4 представляющие собой протонный канал (их может быть от 8 до 16). Точный механизм работы базального тела не известен. Большинство исследователей полагает что поступление протона из периплазмы или внешней среды в MotA4/MotB4 комплекс вызывает конформационные изменения белков, благодаря электростатическому взаимодействию или прямому контакту это изменение приводит к повороту MS-кольца, а его дальнейшее движение возвращает исходную конформацию комплексу и выталкивает протон в цитозоль. У Escherichia coli для одного оборота жгутика требуется перемещение около 1000 протонов. Показано, что жгутик может работать даже у пустых клеточных оболочек при условии что внешний pH ниже внутреннего. Таким образом, базальное тело преобразует химическую энергию в работу, вращаясь за счёт градиента концентрации протонов или, в редких случаях, ионов натрия (некоторые морские бактерии рода Vibrio, алкалофильные Bacillus, Acetobacterium woodii), это вращение осуществляется со скоростью до 100 об/сек, причём его направление может изменяться менее чем за 0,1 сек. Механизм движения клетки: Вращение мотора вызывает пассивное вращение филамента. Более массивная клетка начинает вращаться примерно со скоростью 1/3 от скорости жгутика и в обратном направлении, а также приобретает поступательное движение. Подавляющее большинство наделённых жгутиком бактерий имеют палочковидную форму. Из гидродинамических расчётов следует, что для наиболее эффективного движения отношение длины клетки к ширине должно составлять 3,7. Движение кокков крайне неэффективно, поэтому они чаще всего неподвижны. У ряда бактерий мотор и жгутик могут вращаться только в одном направлении, переориентация происходит при остановке за счёт броуновского движения. Бактерии-перитрихи собирают при движении все свои жгутики (каждый из которых вращается против часовой стрелки) в один пучок. Для совершения кувырка они либо меняют направление вращения, либо изменяют его скорость, из-за чего пучок распадается. При полярном расположении жгутиков один из них может служить и толкающим, и тянущим приспособлением в зависимости от направления вращения. Скорости движения бактерий варьируют от 20 мкм/с у некоторых Bacillus до 200 мкм/с у Vibrio. Особенности жгутиков архей: В результате секвенирования геномов архей не удалось выявить какой-либо гомологии генов, ответственных за биогенез жгутиков архей и бактерий. Вместо флагеллина, неустойчивого в среде с повышенной кислотностью, в жгутиках архебактерий этот белок заменён гликопротеинами. Архебактериальный жгутик тоньше и не имеет центрального полого канала, поэтому, по всей видимости, его синтез происходит по принципиально иному механизму у поверхности клетки. Иначе происходит и вращение мотора жгутика - известно что оно осуществляется.
Cинтез жгутика Процесс синтеза жгутика эубактерий (Caulobacter sp.) запускается экспрессией гена сtrA. Продуктом этого гена является белок CtrA. Синтез Ctr A происходит сразу после перехода клетки из G0-фазы в S-фазу. Обычно участок ДНК, содержащий ген сtrA, метилирован. Синтезу белка CtrA предшествует деметилирование ДНК, которая затем реплицируется. После этого происходит синтез CtrA и его фосфорилирование киназами. Ген сtrA имеет два промотора: Р1 и Р2. С первого промотора осуществляется ингибирование синтеза CtrA. Со второго промотора осуществляется стимулирование синтеза CtrA. Следует отметить, что белок CtrA найден не у всех эубактерий, и механизм синтеза жгутика не универсален. Жгутики эукариот: Жгутики эукариот имеют толщину до 200 нм и длину до 200 мкм. Они окружены выступами цитоплазматической мембраны и содержат 9 пар микротрубочек, выстроенных вокруг двух не объединённых в пару микротрубочек (структура 9+2). Эти микротрубочки скользят друг относительно друга с использованием энергии АТФ, поэтому изгиб эукариотического жгутика может осуществляться в любой его части.
26. Строение, структурная и функциональная классификация синапсов: Синапс - место контакта между двумя нейронами или между нейроном и получающей сигнал эффекторной клеткой. Служит для передачи нервного импульса между двумя клетками, причём в ходе синаптической передачи амплитуда и частота сигнала могут регулироваться. Термин был введён в 1897 г. английским физиологом Чарльзом Шеррингтоном. Структура синапса Типичный синапс - аксо-дендритический химический. Такой синапс состоит из двух частей: пресинаптической, образованной булавовидным расширением окончанием аксона передающей клетки и постсинаптической, представленной контактирующим участком цитолеммы воспринимающей клетки (в данном случае - участком дендрита). Между обеими частями имеется синаптическая щель, края которой укреплены межклеточными контактами. Часть аксолеммы булавовидного расширения, прилежащая к синаптической щели называется пресинаптической мембраной. Участок цитолеммы воспринимающей клетки, ограничивающий синаптическую щель с противоположной стороны, называется постсинаптической мембраной. В синаптическом расширении имеются мелкие везикулы, так называемые синаптические пузырьки, содержащие либо медиатор (вещество-посредник в передаче возбуждения), либо фермент, разрушающий этот медиатор. На постсинаптической и пресинаптической мембранах присутствуют рецепторы к тому или иному медиатору. Классификации синапсов: I. В зависимости от локализации постсинаптической мембраны различают следующие синапсы: 1) аксо-дендритические - с дендритами; 1а) аксо-шипиковые - с дендритными шипиками, выростами на дендритах; 2) аксо-соматические - с телами нейронов; 3) аксо-аксональные - между аксонами; 4) аксо-вазальные - с кровеносной системой (из терминали аксона кроме медиаторов выделяются различные другие секреты, обычно - нейрогормоны); 5) дендро-дендритические - между дендритами; 6) нервно-мышечное окончание - с мышечными волокнами. II. В зависимости от механизма передачи нервного импульса различают: 1) химические; 2) электрические - клетки соединяются плотным контактом с помощью особых коннексонов (каждый из шести белковых субъединиц). Расстояние между мембранами клетки в плотном контакте - 3,5 нм (обычное межклеточное - 20 нм); 3) смешанные синапсы. Наиболее распространён первый тип. III. В зависимости от медиатора синапсы разделяются на: 1) аминергические, содержащие серотонин, дофамин; 1а)адренергические, содержащие адреналин или норадреналин; 2) холинергические, содержащие ацетилхолин; 3) пуринергические, содержащие пурины; 4) пептидергические, содержащие пептиды. При этом в синапсе не всегда вырабатывается только один медиатор. Обычно основной медиатор выбрасывается вместе с другим, играющим роль модулятора. Функционально синапсы можно разделить на: 1) возбудительные - способствуют возникновению возбуждения в постсинаптической клетке (глицинергические (медиатор - глицин) и ГАМК-ергические синапсы (медиатор - гамма-аминомасляная кислота); 2) тормозные - прекращают или предотвращают появление возбуждения в постсинаптической клетке.
В некоторых синапсах присутствует постсинаптическое уплотнение - электронно-плотная зона, состоящая из белков. По её наличию или отсутствию выделяют синапсы асимметричные и симметричные. Известно, что все глутаматергические синапсы асимметричны, а ГАМКергические - симметричны. В случаях, когда с постсинаптической мембраной контактирует несколько синаптических расширений, образуются множественные синапсы. К специальным формам синапсов относятся шипиковые аппараты, в которых с синаптическим расширением контактируют короткие одиночные или множественные выпячивания постсинаптической мембраны дендрита. Шипиковые аппараты значительно увеличивают количество синаптических контактов на нейроне и, следовательно, количество перерабатываемой информации. «Не-шипиковые» синапсы называются «сидячими». Например, сидячими являются все ГАМК-ергические синапсы. Особой формой синапсов являются электротонические, или электрические, в которых благодаря плотному прилеганию синаптической и постсинаптической мембран друг к другу создаются условия для безмедиаторного перехода нервного импульса с цитолеммы одной нервной клетки на другую. Механизм функционирования синапса. При прохождении импульса по синаптической мембране повышается её проницаемость, а из синаптических пузырьков освобождается медиатор, который выходит в синаптическую щель и соединяется с молекулярными рецепторами постсинаптической мембраны, индуцируя на ней возникновение нервного импульса. Медиатор действует в течение очень короткого времени, после чего из синаптических пузырьков освобождается фермент, разрушающий медиатор в синаптической щели. Одновременно часть медиатора перемещается через постсинаптическую мембрану (прямой захват) и в обратном направлении через синаптическую мембрану (обратный захват). Открыты два механизма высвобождения: с полным слиянием везикулы с плазмалеммой и так называемый "поцеловал и убежал", когда везикула соединяется с мембраной, и из неё в синаптическую щель выходят небольшие молекулы, а крупные остаются в везикуле. Второй механизм, предположительно, быстрее первого, по нему происходит синаптическая передача при высоком содержании ионов кальция в синаптической бляшке. Следствием такой структуры синапса является односторонее проведение нервного импульса. Существует так называемая синаптическая задержка - время, нужное для передачи нервного импульса. Её длительность - 0,5 мс. Так называемый «принцип Дейла» (один нейрон - один медиатор) признан ошибочным. Или, как иногда считают, он уточнён: из одного окончания клетки может выделяться не один, а несколько медиаторов, причём этот комплекс постоянен для клетки. История открытия В 1897 году Шеррингтон сформулировал представление о синапсах. За исследования нервной системы, в том числе синаптической передачи, в 1906 году Нобелевскую премию получили Гольджи и Рамон-и-Кахаль. В 1921 австрийский учёный О. Лёви установил химическую природу передачи возбуждения через синапсы и роль в ней ацетилхолина. Получил Нобелевскую премию в 1936 г. совместно с Г. Дейлом. В 1933 советский учёный А. В. Кибяков установил роль адреналина в синаптической передаче. 1970 - Б. Кац (Великобритания), У. фон Эйлер (Швеция) и Дж. Аксельрод (США) получили Нобелевскую премию за открытие роли норадреналина в синаптической передаче.
27. Непостоянные включения в клетке: Включения клетки - все структуры цитоплазмы клетки. Обычно кключения клетки подразделяют на 3 группы: 1) постоянные, или органоиды, осуществляющие общие функции клетки (митохондрии, Гольджи комплекс, хлоропласты); 2) временные, или параплазматические, образования, появляющиеся и исчезающие в процессе обмена веществ (секреторные гранулы, питательные вещества, жир, крахмал); 3) специальные, или метаплазматические, образования, имеющиеся в некоторых специализированных клетках, где они выполняют частные функции, например сокращения (миофибриллы мышечных клеток), опоры (тонофибриллы в клетках эпидермиса). Гранулы секреторные - округлые гранулы, появляющиеся и исчезающие в цитоплазме в процессе нормального обмена веществ. Ультраструктура функциональных клеток передней доли гипофиза крысы (в норме):
 Описание
рисунка: Соматотрофы (1) с выраженной
эндоплазматической сетью и секреторными
гранулами (2); лактотроф (3) с крупными
секреторными гранулами (2); кортикотрофы
(4) с мелкими секреторными гранулами.
Гранулы
тучных клеток цитоплазматические.
В
цитоплазматических гранулах тучных
клеток находятся медиаторы и модуляторы
воспаления, пролиферации и миграции
клеток (гистамин, нейтральные протеазы
- химаза и триптаза, кислые гидролазы,
катепсин G, карбоксипептидаза,
гепарин-сульфат, хондроитин-сульфат,
протеогликаны), а также медиаторы
воспаления, которые синтезируются при
активации клетки (простагландин D2,
лейкотриен C4, цитокины, воспалительный
белок макрофагов).
Хромафинные
гранулы.
В
мозговом слое надпочечников содержатся
хромафинные гранулы - органеллы, способные
к биосинтезу, поглощению, запасанию и
секреции катехоламинов. Свое название
они получили потому, что окрашиваются
под действием бихромата калия в
красно-коричневый цвет. Помимо
катехоламинов в состав этих гранул
входит ряд других веществ, в том числе
и белок хромогранин А.
Гранулы
плотные тромбоцитов.
В
плотных гранулах накапливаются и
хранятся неметаболический пул, серотонин
и ионы кальция. В альфа-гранулах содержатся
фактор 4 тромбоцитов, бета-тромбоглобулин,
тромбоспондин, фибронектин, тромбоцитный
фибриноген, тромбоцитный фактор
Виллебранда, факторы роста и другие
белки.
Гранулы-альфа
тромбоцитов.
В
альфа-гранулах содержатся фактор 4
тромбоцитов, бета-тромбоглобулин,
тромбоспондин, фибронектин, тромбоцитный
фибриноген, тромбоцитный фактор
Виллебранда, факторы роста и другие
белки.
Гранулы
ацидофильные эозинофилов.
Эозинофилы
содержат крупные овальные ацидофильные
гранулы, состоящие из аминокислот,
белков и липидов. Главный основный белок
локализован в ядре гранул, в то время
как катионный белок эозинофилов и
пероксидаза находятся в матриксе гранул.
Арилсульфатаза B, фосфолипаза D и
гистаминаза также включены в гранулы.
Реакция дегрануляции - это один из
механизмов использования эозинофилами
токсичного содержимого своих гранул.
Описание
рисунка: Соматотрофы (1) с выраженной
эндоплазматической сетью и секреторными
гранулами (2); лактотроф (3) с крупными
секреторными гранулами (2); кортикотрофы
(4) с мелкими секреторными гранулами.
Гранулы
тучных клеток цитоплазматические.
В
цитоплазматических гранулах тучных
клеток находятся медиаторы и модуляторы
воспаления, пролиферации и миграции
клеток (гистамин, нейтральные протеазы
- химаза и триптаза, кислые гидролазы,
катепсин G, карбоксипептидаза,
гепарин-сульфат, хондроитин-сульфат,
протеогликаны), а также медиаторы
воспаления, которые синтезируются при
активации клетки (простагландин D2,
лейкотриен C4, цитокины, воспалительный
белок макрофагов).
Хромафинные
гранулы.
В
мозговом слое надпочечников содержатся
хромафинные гранулы - органеллы, способные
к биосинтезу, поглощению, запасанию и
секреции катехоламинов. Свое название
они получили потому, что окрашиваются
под действием бихромата калия в
красно-коричневый цвет. Помимо
катехоламинов в состав этих гранул
входит ряд других веществ, в том числе
и белок хромогранин А.
Гранулы
плотные тромбоцитов.
В
плотных гранулах накапливаются и
хранятся неметаболический пул, серотонин
и ионы кальция. В альфа-гранулах содержатся
фактор 4 тромбоцитов, бета-тромбоглобулин,
тромбоспондин, фибронектин, тромбоцитный
фибриноген, тромбоцитный фактор
Виллебранда, факторы роста и другие
белки.
Гранулы-альфа
тромбоцитов.
В
альфа-гранулах содержатся фактор 4
тромбоцитов, бета-тромбоглобулин,
тромбоспондин, фибронектин, тромбоцитный
фибриноген, тромбоцитный фактор
Виллебранда, факторы роста и другие
белки.
Гранулы
ацидофильные эозинофилов.
Эозинофилы
содержат крупные овальные ацидофильные
гранулы, состоящие из аминокислот,
белков и липидов. Главный основный белок
локализован в ядре гранул, в то время
как катионный белок эозинофилов и
пероксидаза находятся в матриксе гранул.
Арилсульфатаза B, фосфолипаза D и
гистаминаза также включены в гранулы.
Реакция дегрануляции - это один из
механизмов использования эозинофилами
токсичного содержимого своих гранул.
Питательные вещества: 1) Белки – основной строительный материал мышечной и костной ткани. Белки входят в состав большинства ферментов (пепсин, трипсин, амилаза) и гормонов. Транспортная функция белков обеспечивает перенос с кровью различных веществ к тканям (кислорода, жиров). Защитная функция белков особого типа (иммуноглобулинов) обеспечивает иммунитет, способ защиты внутреннего постоянства организма от живых тел и веществ, несущих в себе признаки генетически чужеродной информации. 2) Жиры. К жирам (липидам) относят большую группу содержащихся в живых клетках органических веществ с различным химическим строением и некоторыми общими физико-химическими свойствами. Такими общими свойствами липидов являются их нерастворимость в воде (гидрофобность) и растворимость в неполярных растворителях: ацетоне, спиртах, бензоле. С точки зрения состава, под жирами подразумевают строго определенные соединения сложных эфиров высших жирных кислот и спиртов. Жиры входят в состав тканей; в больших количествах они содержатся в головном и спинном мозге, печени, сердце, а их концентрация в нервной ткани достигает 25%. 3) Углеводы – основной источник энергии для всего организма в целом. Углеводы выполняют в организме и ряд специализированных функций, связанных со специфичностью группы крови и ее сворачиваемостью в сосудах. Углеводы подразделяют на три основных класса: - моносахариды (глюкоза и фруктоза); - дисахариды (сахароза); - полисахариды (крахмал и гликоген). 4) Пищевые волокна – волокна активно влияющие на обмен веществ и необходимые для нормальной жизнедеятельности организма. Пищевые волокна обладают следующими свойствами: - связывают воду, что приводит к их набуханию; - абсорбируют (поглощают) токсичные вещества и выводят их из организма; - снижают уровень холестерина; - усиливают раздражающее действие пищи, что приводит к стимулированию перистальтики кишечника и более быстрому транзиту пищи; - нормализуют полезную микрофлору кишечника. Если пищевые волокна в избытке, то они связывают и удаляют из организма не только шлаки, но и часть полезных компонентов пищи.
5) Витамины – это группа низкомолекулярных незаменимых факторов, которые обладают выраженной биологической активностью. Витамины способны улучшать внутреннюю среду, повышать устойчивость организма к неблагоприятным факторам, повышать работоспособность, замедлять процессы старения и являться надежным средством общей профилактики болезней. Все они имеют большое значение в регуляции обмена веществ и физиологических функций. -Витамин А (ретинол) необходим для здоровой кожи, волос, зубов и ногтей, процесса роста, обеспечения нормального зрения, а также для усвоения белка организмом. Повышает сопротивляемость респираторным инфекциям. - Витамин D поддерживают нормальное состояние костных тканей в организме. Необходим для крепких костей и зубов. - Витамин Е обладает ярко выраженным антиоксидантным действием. Уменьшает воздействие свободных радикалов, разрушающе действующих на клетки. Улучшает снабжение организма кислородом. - Витамин B1 имеет важное значение в углеводном, белковом и жировом обмене. Жизненно необходим для здоровой нервной системы. - Витамин В2 входит в состав ферментов, участвующих в процессе переработки белков, жиров и углеводов. Помогает клеткам выводить продукты распада. - Витамин B3 участвует в расщеплении углеводов. Необходим для синтеза натуральных половых гормонов и инсулина. Важен для циркуляции, переноса и поглощения кислорода клетками. - Витамин В6 играет важную роль в белковом и жировом обмене, а также в регуляции кислотности и желудочной секреции. Необходим для нормального функционирования и роста красных кровяных телец и антител. - Витамин РР (никотиновая кислота, витамин В5) участвует в функционировании большого количества ферментов. Необходим для синтеза натуральных половых гормонов. Важен для циркуляции и переноса кислорода клетками. - Витамин В12 участвует в синтезе аминокислот. Способствует переработке жировых отложений в энергию. - Фолиевая кислота участвует в метаболизме белков и углеводов, способствует выделению желудком соляной кислоты для эффективного переваривания белка. Необходима для формирования и воспроизводства клеток. - Витамин С (аскорбиновая кислота) усиливает действие всех вышеперечисленных витаминов. Необходим для нормального белкового обмена, для образования соединительной ткани, в том числе в стенках кровеносных сосудов, для синтеза стероидных гормонов, играющих важную роль в адаптации организма при стрессовых ситуациях и регулирующих иммунитет.