

Питання 3. Нейроендокринна регуляція линьки хордових
Процеси линьки активуються гіпоталамо-гіпофізарно-тиреоїдною системою. Гіпоталамус виділяє тиреоліберин, який активує синтез тиреоліберину гіпофізом. Тиреоліберин активує синтез тиреоїдних гормонів щитовидною залозою. Гормони щитовидної залози активують линьку.
Час настання сезонної та шлюбної линьки визначається довжиною світлового дня (фотоперіодом). У сприйнятті довжини світлового дня задіяний епіфіз та його гормон мелатонін. Відомо, що існує добовий ритм секреції мелатоніну: багато цього гормона утворюється у темний період доби, і мало – у світлий період доби. Взимку, коли ночі довгі, мелатоніну за добу зрозуміло утвориться більше, ніж влітку, коли ночі короткі. Таким чином, середньодобовий рівень мелатоніну корелює із порою року. Линька запускається при певному середньодобовому рівні мелатоніну.
Існує також зворотня кореляція між ступенем активності гонад та інтенсивністю линьки. Коли гонади активні, линька не відбувається. І навпаки, коли відбувається линька, гонади неактивні. Очевидно, це необхідно для того, щоб розмежувати в часі сезон розмноження та процеси линьки, оскільки обидва процеси вимагають великих витрат пластичних та енергетичних ресурсів організму.
Регулюється линька у ссавців так само, як і у птахів. Вона активується гормонами щитовидної залози. Сезонність линьки регулюється мелатоніном епіфіза, а зовнішнім регулятором є довжина світлового дня. На процеси линьки у ссавців сильний вплив має також температура. Наприклад, у багатьох видів ссавців довжина та щільність розташування волосся різні у представників різних популяцій, що мешкають в різних природно-кліматичних умовах.
Питання 4. Гормональна регуляція росту.
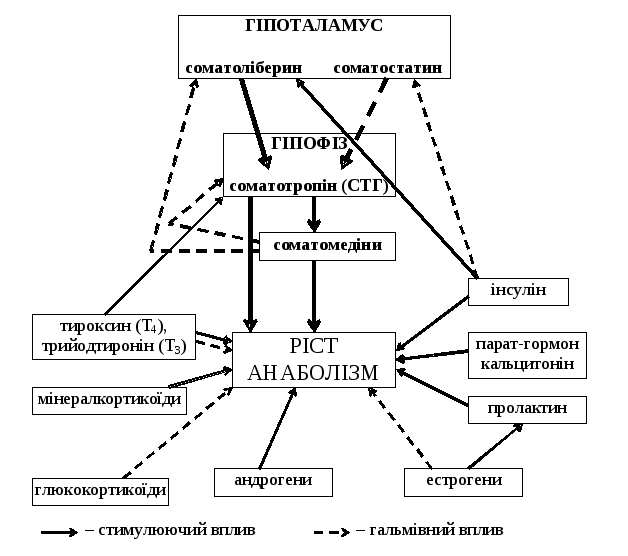
Ріст у хребетних регулюється, в основному, гіпоталамо-гіпофізарною системою (мал. 11).
Основним є гормон аденогіпофіза – соматотропін (СТГ), який стимулює процеси росту організму. Причому, у ростучих тканинах спостерігаються і анаболічний, і проліферативний ефекти, а у зрілих тканинах – майже тільки анаболічний.
 Мал.
11. Нейроендокринна регуляція росту у
хребетних.
Мал.
11. Нейроендокринна регуляція росту у
хребетних.
До соматотропіну найбільш чутлива хрящова тканина, особливо, епіфізи трубчастих кісток. Соматотропін тут стимулює хондрогенез (ріст хряща), тобто, проліферацію хрящових клітин та синтез білків (колагену, мукополісахаридів). Внаслідок цього трубчасті кістки ростуть у довжину. А оскільки ріст всього тіла залежить від росту скелету, то відбувається ріст всього організму. Після закриття епіфізарних хрящів ця зона стає нечутливою до дії соматотропіну.
Соматотропін у власне сполучній тканині посилює проліферацію фібробластів (сприяє їх переходу із G1‑фази в S‑фазу), стимулює синтез колагену та хондроітинсульфату (основного компонента міжклітинної аморфної речовини). Соматотропін стимулює також проліферацію клітин та біосинтез білка у внутрішніх паренхіматозних органах. У тимусі соматотропін посилює імунопоез і стимулює синтез тимозинів. Стимуляція біосинтезу білка відбувається у три стадії: спочатку – посилення транспорту в клітину через плазмалему амінокислот та глюкози (відбувається краще забезпечення клітини будівельним матеріалом для синтезу білка), потім – прискорення трансляції на рибосомах, і нарешті – посилення синтезу рРНК.
Соматотропін окрім прямої дії, стимулює синтез соматомедінів у печінці та деяких інших органах, які в подальшому стимулюють процеси росту певних тканин. До соматомедінів можна віднести: соматомедін С (інсуліноподібний фактор росту І), фактор росту нервів, фактор росту тромбоцитів, фактор росту епідермісу, тимозини, простагландини. За принципом негативного зворотнього зв’язку соматомедін С інгібує синтез соматоліберину і соматотропіну.
Окрім системи соматотропін–соматомедіни, яка є головною в регуляції процесів росту, на ріст організму впливає і ряд інших гормонів: інсулін, мінералкортикоїди, глюкокортикоїди, тиреоїдні гормони, андрогени, естрогени, пролактин, парат‑гормон, кальцитонін (мал. 11). Інсулін діє подібно до соматотропіну. Він стимулює біосинтез білка в м’язах, печінці, сполучній тканині; проте на хрящову тканину дія слабка. Інсулін стимулює синтез соматоліберину і пригнічує синтез соматостатину гіпоталамусом. Тиреоїдні гормони (тироксин та трийодтиронін) в низьких дозах активують, а у високих дозах – пригнічують ріст. Ця їх дія, мабуть, опосередковується через стимулюючий вплив на інтенсивність метаболізму (невелике посилення метаболізму сприяє більш швидкому росту, а надмірне посилення метаболізму, коли падає ефективність використання поживних речовин, гальмує ріст). При дії тиреоїдних гормонів у малих дозах відбувається стимуляція біосинтезу білка на рівні транскрипції (у м’язах, печінці, але не у хрящах), а при їх дії у великих дозах – гальмування біосинтезу білка (через роз’єднання процесів окислення і фосфорилювання у мітохондріях, внаслідок чого створюється дефіцит АТФ). Тиреоїдні гормони також стимулюють соматотропін-продукуючі клітини в аденогіпофізі. Андрогени, головним чином, тестостерон, стимулює анаболізм, що веде до інтенсифікації процесів росту, особливо, під час статевого дозрівання. Вони стимулюють ріст м’язів, органів чоловічої статевої системи. Але на тимус андрогени діють катаболічно, а тому він інволює під час статевого дозрівання. Естрогени, головним чином, естрадіол, стимулюють закриття епіфізарних хрящів, що веде до припинення росту трубчастих кісток у довжину, і, як наслідок, до припинення росту організму. Естрогени стимулюють ріст м’язів, печінки, органів жіночої статевої системи. Пролактин у нижчих хребетних нарівні із соматотропним гормоном активує процеси росту організму. У птахів та ссавців пролактин грає меншу роль; він активує ріст волосся, пір’я, молочних залоз, деяких внутрішніх органів. Глюкокортикоїди, головним чином, кортизол, мають катаболічний ефект (гальмують біосинтез білка на рівні транскрипції), а тому гальмують ріст організму. Але в печінці вони мають анаболічний ефект. Мінералкортикоїди, головним чином, альдостерон, активують Na+‑K+‑АТФазу, це веде до зростання концентрації K+ в клітині, що дає змогу діяти соматотропному гормонові, соматомедінам та інсулінові. Парат‑гормон та кальцитонін регулюють обмін Ca2+, а тому мають вплив на ріст скелету.