

Список продуктов содержащих глутамат натрия
«Магги» бульонные кубики, ЗАО Нестле.Икорная закуска, ООО Полезные продукты.Говядина тушёная, ОАО «Дейма».«Роллтон», ЗАО Ди Эч Ви-е.«Биг Ланч» лапша быстрого приготовления, ООО Биофуд.«Хрустим» сухарики, ООО «Фритолеймануфактуринг».«Лейс» чипсы, ООО «Фритолеймануфактуринг». «Читос» чипсы, ООО «Фритолеймануфактуринг».«Русская картошка» чипс, ООО Рускарт. «Доширак», ООО Коя Раменское. «Кнорр» приправы ООО «Юнилевер». «Рыбка к пиву», ООО Моридо. «Воронцовские сухарики», ООО РУСКО.«Кальмар сушёный», ООО Моридо. «Кириешки», Сибирский берег. «Роллтон» домашний бульон-ЗАО Ди Эч Ви-С«Биг Ланч» картофельное пюре, ООО Биофуд. Суп ООО «Приправыч». Тушенка с говядиной кусковая, Еленский пищевой комбинат.Котлеты ООО «Инвест Альянс» Ермолино.«Кордон Блю» ООО «Инвест Альянс» Ермолино.«Голубцы» ООО «Инвест Альянс» Ермолино.«Чебуреки» ООО «Инвест Альянс» Ермолино.«Шницель особенный» ООО «Инвест Альянс» Ермолино.«Пельмени русские» ООО «Инвест Альянс» Ермолино.«Пельмени домашние» ООО «Инвест Альянс» Ермолино. «Блинчики с мясом» ООО «Инвест Альянс» Ермолино.
Взаимосвязь глутамата и глутаматергич нейромедиаторной системы
Возбуждающее действие глутамата известно с 1950-х .Различают два подтипа рецепторов, через которые глутамат опосредует свои регуляторные влияния: ионотропные и метаботропные. Активация ионотропных рецепторов обеспечивает формирование потенциала действия, тогда как метаботропные регулируют его величину и длительность .
Ионотропные рецепторы глутамата играют значительную роль в возбуждающей синаптической передаче в ЦНС млекопитающих. Различные подтипы этих рецепторов обнаруживаются более чем в 50 % синаптических контактов ЦНС. На основании фармакологических исследований ионотропные рецепторы глутамата были сгруппированы в три различных подкласса: AMPA, КА и NMDA-рецепторы. Эти рецепторы содержат ионные каналы, проницаемые катионами, относительная проницаемость для кальция и натрия колеблется в зависимости от подкласа и субъединичной композиции.
Метаботропные рецепторы глутамата ассоциированы с G-белками, они также подразделяются на подклассы в зависимости от эффекторной системы.Помимо глутамата другими эндогенными лигандами ионотропных рецепторов глутамата являются : L-аспартат ; сульфоновые и сульфиновые аналоги глутамата и аспартата (т.е. L-цистеин сульфонат, L-гомоцистеин сульфонат, L-гомоцистеат, L-цистеат). Хинолинат, выделяемый астроцитами – селективный и слабый агонист NMDA рецепторов, вызывающий нейродегенегацию в гиппокампе и стриатуме при фокальной инъекции. Эндогенный дипептид, N-ацетил-аспартил-глутамат, (NAAG) агонист НМДА в латеральных коленчатых ядрах. Для активации NMDA рецепторов требуется глицин или D-серин в качестве ко-агониста. Выброс глутамата из везикул пресинаптических терминалей осуществляется по кальций-зависимому механизму, в котром участвуют потенциал-зависимые кальциевые каналы N- и P/Q-типа . Выброс глутамата может также осуществляться и при обратной работе транспортеров глутамата. Это происходит, когда при церебральной ишемии снижаются градиенты калия и натрия . Синаптический выброс глутамата контолируется различными пресинаптическими рецепторами, включая не только метаботропные рецепторы глутамата 2 и 3 класса, но и М- и Н-ацетилхолиновые, А1-аденозиновые, каппа-опиоидные, ГАМКб-рецепторы, рецепторы холецистокинина и нейропептида Y (Y2). Из ЦНС млекопитающих было клонированно пять переносчиков глутамата. Два из них экспрессируются преимущественно в глии (глиальный транспортер глутамата и аспартата - GLAST, и глиальный транспортер глутамата - GLT) и три в нейронах [, транспортеры возбуждающих аминокислот EAAC1 ,EAAT4 и EAAT5] (у человека они называются, соответственно, EAAT1–5). Эти транспортеры натрий-зависимы, градиент натрия и калия является источником энергии для этого транспорта.
Глутаматергические и аспартатергические нейроны Структурно сходные аминокислоты глутамат и аспартат электрофизиологически классифицируются как возбуждающие нейротрансмиттеры. Нервные клетки, содержащие глутамат и/или аспартат в качестве нейротрансмиттеров, имеются в слуховой системе (нейроны первого порядка), в обонятельной системе (объединяют обонятельную луковицу с корой большого мозга), в лимбической системе, в неокортексе (пирамидные клетки). Глутамат обнаруживается также в нейронах проводящих путей, идущих от пирамидных клеток: кортикостриарном, кортикоталамическом, кортикотектальном, кортикомостовом и кортикоспинальном трактах.
Важную роль в функционировании глутаматной системы играют астроциты, не являющиеся пассивными элементами нервной системы, а участвующие в обеспечении нейронов энергетическими субстратами в ответ на увеличение синаптической активности. Астроцитарные отрост-
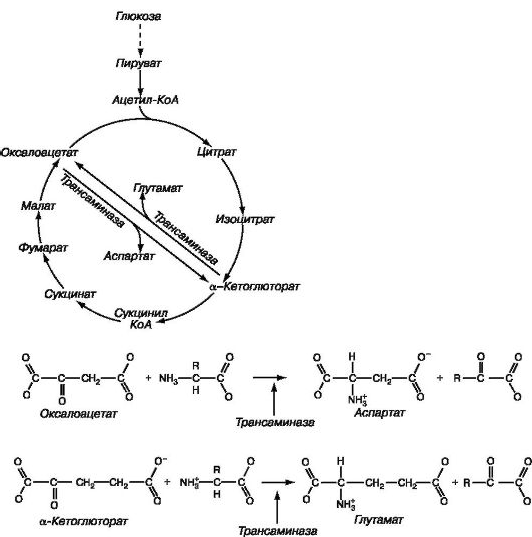
Рис. 1.34. Синтез глутаминовой и аспарагиновой кислот.
Путем гликолиза происходит превращение глюкозы в пируват, который в присутствии ацетил-КоА вступает в цикл Кребса. Далее путем трансаминирования оксалоацетат и α-кетоглутарат превращаются в аспартат и глутамат соответственно (реакции представлены в нижней части рисунка)
ки расположены вокруг синаптических контактов, что позволяет им улавливать увеличение синаптической концентрации нейротрансмиттеров (рис. 1.35). Перенос глутамата из синаптической щели опосредуется специфическими транспортными системами, две из которых глиальноспецифичны (GLT- 1 и GLAST- переносчики). Третья транспортная система (ЕААС-1), находящаяся исключительно в нейронах, не вовлекается в перенос высвободившегося из синапсов глутамата. Переход глутамата в астроциты происходит по электрохимическому градиенту ионов Na+.
В нормальных условиях поддерживается относительное постоянство внеклеточных концентраций глутамата и аспартата. Их повышение включает компенсаторные механизмы: захват нейронами и астроцитами избытков из межклеточного пространства, пресинаптическое торможение выброса нейротрансмиттеров, метаболическую утилизацию и

Глутамат высвобождается из синаптических везикул в синаптическую щель. На рисунке изображены два механизма обратного захвата: 1 - обратно в пресинаптическое окончание; 2 - в соседнюю глиальную клетку; 3 - глиальная клетка; 4 - аксон; 5 - глутамин; 6 - глутамин-синтетаза; 7 - АТФ+NH4+; 8 - глутаминаза; 9 - глутамат + NH4+; 10 - глутамат; 11 - постсинаптическая мембрана. В глиальных клетках глутаминсинтаза превращает глутамат в глутамин, который далее переходит в пресинаптическое окончание. В пресинаптическом окончании глутамин превращается обратно в глутамат ферментом глутаминазой. Свободный глутамат также синтезируется в реакциях цикла Кребса в митохондриях. Свободный глутамат собирается в синаптических везикулах до возникновения следующего потенциала действия. В правой части рисунка представлены реакции превращения глутамата и глутамина, опосредованные глутаминсинтетазой и глутаминазой
др. При нарушении их элиминации из синаптической щели абсолютная концентрация и время пребывания глутамата и аспартата в синаптической щели превышают допустимые пределы, и процесс деполяризации мембран нейронов становится необратимым.
Ионотропные рецепторы играют ключевую роль в осуществлении возбуждающей нейропередачи в ЦНС, реализации нейропластичности, образовании новых синапсов (синаптогенез), в повышении эффективности функционирования уже имеющихся синапсов. С указанными процессами во многом связаны механизмы памяти, обучения (приобретение новых навыков), компенсации функций, нарушенных вследствие органического поражения мозга.
Имеются данные о нейротоксических свойствах глутамата, связанных с активацией АМРА- и К-рецепторов, приводящей к изменению проницаемости постсинаптической мембраны для одновалентных катионов К+ и Na+, усилению входящего тока ионов Na+ и кратковременной деполяризации постсинаптической мембраны, что, в свою очередь, вызывает усиление притока Са2+ в клетку через агонист-зависимые (рецепторы NMDA) и потенциалзависимые каналы. Поток ионов Na+ сопровождается входом в клетки воды, что обусловливает набухание апикальных дендритов и лизис нейронов (осмолитическое повреждение нейронов).
Метаботропные глутаматные рецепторы, связанные с G-белком, играют важную роль в регуляции внутриклеточного кальциевого тока, вызванного активацией NMDA-рецепторов, и выполняют модуляторные функции, вызывая тем самым изменения в деятельности клетки. Эти рецепторы не влияют на функционирование ионных каналов, но стимулируют образование внутриклеточных посредников диацилглицерола и нозитолтрифосфата, принимающих участие в дальнейших процессах ишемического каскада.