

Пути утилизации кислорода клеткой
Большая часть кислорода, потребляемого клеткой (около 80 %), используется, как указано, в митохондриях с участием цитохромоксидазы. Это так называемый оксидазный путь. При этом происходит полное восстановление кислорода, причем субстрат не реагирует с кислородом непосредственно. Данный путь дает клетке энергию в виде АТФ. Помимо цитохромоксидазы существуют другие оксидазы (ФМН и ФАД-зависимые), которые катализируют реакции окисления веществ с образованием перекиси водорода.
Н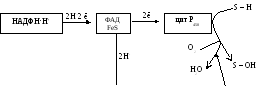 аряду
с этим существует другой путь окисления
—оксигеназный. Он не дает клетке
энергии, кислород включается в субстрат
с образованием новой гидроксильной или
карбоксильной группы. Этот путь происходит
в основном в мембранах эндоплазматического
ретикулума(микросомах). Путеммикросомного окисленияосуществляется
α- и ω-окисление жирных кислот, синтез
ненасыщенных жирных кислот, стероидов.
Таким путем обезвреживаются ксенобиотики,
т. е. чужеродные для организма вещества
(лекарства, ядохимикаты, косметические
препараты). Ферменты, осуществляющие
такое окисление, называются оксигеназами.Различают диоксигеназы,которые
включают в молекулу субстрата два атома
молекулы кислорода. Более распространены
в клетках монооксигеназы (гидроксилазы).
Они катализируют реакции, при которых
в молекулу субстрата включается один
атом из молекулы кислорода, второй же
атом кислорода восстанавливается при
этом до воды. Монооксигеназные системы
представляют собой короткие цепи
переноса электронов и протонов, источником
которых служит чаще всего восстановленный
НАДФ+, реже НАД+или аскорбиновая
кислота. Активатором кислорода при этом
являетсяцитохром Р450—
одноцепочечный хромопротеин с молекулярной
массой 50 кДа. Примерная схема
монооксигеназной цепи представлена на
рисунке 7.4.
аряду
с этим существует другой путь окисления
—оксигеназный. Он не дает клетке
энергии, кислород включается в субстрат
с образованием новой гидроксильной или
карбоксильной группы. Этот путь происходит
в основном в мембранах эндоплазматического
ретикулума(микросомах). Путеммикросомного окисленияосуществляется
α- и ω-окисление жирных кислот, синтез
ненасыщенных жирных кислот, стероидов.
Таким путем обезвреживаются ксенобиотики,
т. е. чужеродные для организма вещества
(лекарства, ядохимикаты, косметические
препараты). Ферменты, осуществляющие
такое окисление, называются оксигеназами.Различают диоксигеназы,которые
включают в молекулу субстрата два атома
молекулы кислорода. Более распространены
в клетках монооксигеназы (гидроксилазы).
Они катализируют реакции, при которых
в молекулу субстрата включается один
атом из молекулы кислорода, второй же
атом кислорода восстанавливается при
этом до воды. Монооксигеназные системы
представляют собой короткие цепи
переноса электронов и протонов, источником
которых служит чаще всего восстановленный
НАДФ+, реже НАД+или аскорбиновая
кислота. Активатором кислорода при этом
являетсяцитохром Р450—
одноцепочечный хромопротеин с молекулярной
массой 50 кДа. Примерная схема
монооксигеназной цепи представлена на
рисунке 7.4.
Рис. 7.4. Механизм микросомного окисления
Смысл такого процесса заключается в том, что ксенобиотики, которые обычно гидрофобны, гидроксилируясь, становятся более гидрофильными, что способствует их обезвреживанию и выведению из организма с желчью или мочой. С участием микросомных систем осуществляется также биосинтез стероидов, желчных кислот, витамина Д3.
С появлением в атмосфере кислорода, а он появился тогда, когда возникли фотосинтезирующие организмы, стало возможным более эффективно использовать энергию, т. е. возник механизм окислительного фосфорилирования. Но, с другой стороны, вместе с этим кислород принес и новую опасность. При неполном восстановлении молекулы кислорода образуются высокоактивные формы (свободные радикалы),которые могут повреждать белки, нуклеиновые кислоты, липиды и способны даже убить живую клетку. Активные формы кислорода или свободные радикалы образуются в качестве промежуточных продуктов в ходе микросомного окисления, при работе дыхательной цепи, при воздействии ионизирующего излучения, при самопроизвольном окислении ряда веществ (гемоглобин).Свободные радикалы— молекулы, содержащие неспаренные электроны, агрессивные молекулы, которые атакуют другие молекулы с целью отнять у них электрон. К ним относятся: супероксидный анион-радикал (О2·–), гидропероксидный радикал (НО2·), пероксид водорода (Н2О2), гидроксидный радикал (НО·).
Свободные радикалы стимулируют разрывы в молекулах нуклеиновых кислот, нарушают функции белков, ведут к деполимеризации протеогликанов соединительной ткани, повреждают ненасыщенные жирные кислоты клеточных мембран, запуская, тем самым, механизм перекисного окисления липидов (ПОЛ). Вместе с тем свободные радикалы кислорода играют и положительную роль, так как участвуют в осуществлении фагоцитами и Т-лимфоцитами их защитной функции.
Повышенное образование в организме свободных радикалов кислорода ведет к «окислительному стрессу», который может привести к повреждению мембран и гибели клетки. Поэтому в организме существует антиоксидантная защита от свободных радикалов.
Различают неферментативную и ферментативную защиту клеток. Важнейшим компонентом неферментативной защиты является витамин Е (токоферол), витамин размножения.
Являясь жирорастворимым витамином, он всасывается вместе с липидами, поступает в лимфатическую систему и кровяное русло, а оттуда — в ткани. Токоферол защищает ненасыщенные жирные кислоты клеточных мембран от перекисного окисления, предохраняет от окисления SH-группы мембранных белков, защищает от окисления двойные связи в молекулах каротинов и витамина А. Токоферол (совместно с витамином С) способствует включению селена в состав активного центра глутатионпероксидазы — важнейшего фермента антиоксидантной защиты клеток. Он контролирует синтез гема, цитохромов, стабилизирует биологические мембраны.
Ферментативная защита клеток от свободных радикалов (антиоксидантная защита) осуществляется с помощью следующих ферментов:
супероксиддисмутазы(превращает супероксидные радикалы в менее токсичную перекись водорода);
каталазы(разлагает перекись водорода на воду и кислород);
глутатионпероксидазы— главная система защиты эритроцитов от разрушительного действия перекиси водорода. В качестве кофермента глутатионпероксидаза использует трипептид – глутатион. Для ее работы также необходим микроэлемент селен (Se).