

Тема 13. Внутриклеточный метаболизм жирных кислот
Жирные кислоты проходят через клеточную мембрану путем диффузии по концентрационному градиенту.
Активация жирных кислот. Первым этапом на пути метаболизма длинноцепочечных жирных кислот в клетке является их активация за счет образования ацил-КоА. Эту реакцию катализирует фермент ацил-КоА синтетаза, который локализован на наружной мембране митохондрий:
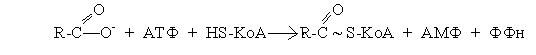
Перенос ацил-КоА в митохондрии. Ацил-КоА, имеющий средней длины или короткую углеводородную цепь (<10), может проходить через митохондриальную мембрану путем диффузии (рис. 13.1). Перенос длинноцепочечного ацил-КоА происходит с помощью карнитин-ацил- трансфераз (КАТ), локализованных соответственно на наружной и внутренней мембранах митоходрий, и транслоказы (Т). Предшественник для процесса синтеза жирных кислот, малонил-КоА, является аллостерическим ингибитором активности КАТ.
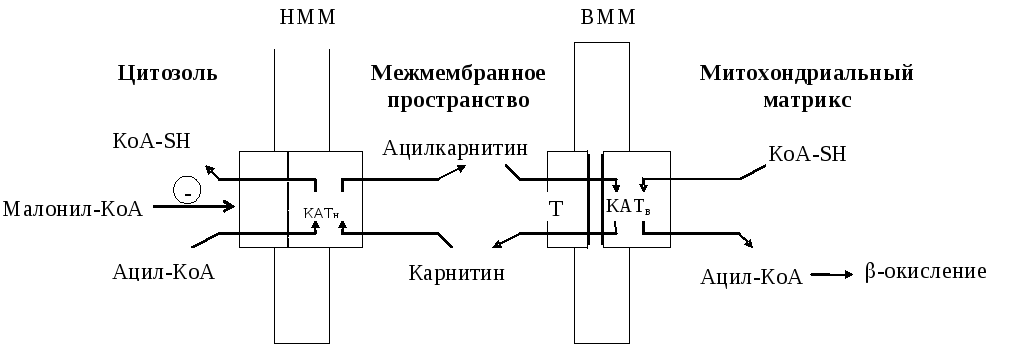
Рис. 13.1. Перенос ацил-КоА в митохондрии
-ОКИСЛЕНИЕ АЦИЛ-КоА
Последовательность реакций -окисления катализируется 4 ферментами. С их помощью идет дегидрирование, гидратация, образование -кетокислоты и тиолитическое расщепление с высвобождением двухуглеродных фрагментов (ацетил-КоА) (рис. 13.2).
Энергетический выход -окисления на примере пальмитиновой кислоты. Образование АТФ (1,5 АТФ/ФАДН2; 2,5 АТФ/НАДН.H+; 10 АТФ/ацетил-КоА; таким образом, для пальмитоил-КоА (жирная кислота с 16 С): 7 ФАДН2, 7 НАДН.H+ и 8 ацетил-КоА = 108 АТФ).
Расход АТФ на активацию — 2 АТФ (используется энергия гидролиза двух макроэргических связей), в ходе которой пальмитат превращается в пальмитоил-КоА. Чистый энергетический выход для окисления пальмитата — 106 АТФ.
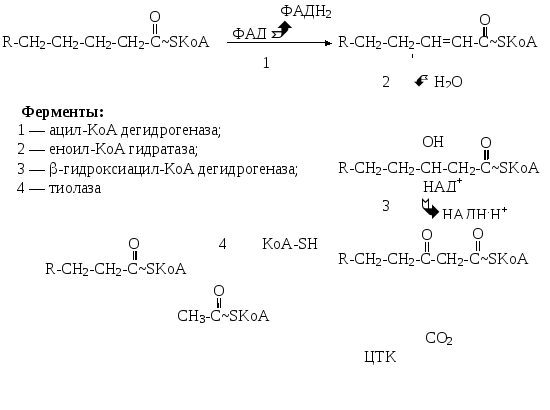
Рис. 13.2. Реакции -окисления жирных кислот
Окисление ненасыщенных жирных кислот. В ходе -окисления ненасыщенных жирных кислот отщепление двухуглеродных фрагментов ведет к образованию ацил-КоА с двойной связью в цис-положении между С3 и С4. Затем с помощью фермента еноил-КоА изомеразы происходит её превращение в транс-двойную связь между С-2 и С-3. Другой фермент, 2,3-диеноил-КоА редуктаза, может катализировать насыщение двойной связи между С-4 и С-5 в составе ацил-КоА с использованием в качестве кофермента НАДН.Н+. Образовавшийся промежуточный продукт подвергается дальнейшему превращению под влиянием еноил-КоА изомеразы.
Окисление жирных кислот в пероксисомах
Окисление жирных кислот в пероксисомах составляет около 30 % всего их окисления. В пероксисомах окисляются необычные жирные кислоты (с длинной углеводородной цепью, дикарбоновые, с разветвленным радикалом). Укорочение радикала в пероксисомах происходит до тех пор, пока не образуется ацил-КоА со средней длиной цепи. Образующийся ацил-КоА с С-8 впоследствии подвергается дальнейшему окислению в митохондриях.
Первоначальная стадия дегидрирования в ходе пероксисомного окисления жирных кислот протекает с образованием Н2О2, а не ФАДН2. Перекись водорода удаляется с помощью каталазы. Все последующие реакции аналогичны происходящим в митохондриях, хотя катализируются они изоферментами пероксисом.