

Результаты и обсуждение
В 1-й серии в полость желудка (ПЖ) при помощи зонда вводили 1 мл 1% раствора МЧ, а во 2-й серии -такое же количество МЧ в/б. В контроле (К) вводили по
1 мл изотонического раствора - 0,9 % NaCI (ИР). В обеих сериях начало наступления анестезии и её продолжительность не отличались от контроля.
В 3-й и 4-й сериях стимуляцию секреции ЖК вызывали путем введения в ПЖ 1 мл оливкового масла (К -1 мл ИР), на фоне которой изучали анестезирующее действие лидокаина и скандонеста, получены аналогичные результаты. В них четко прослеживается укорочение продолжительности действия препаратов, но у скандонеста оно было более продолжительным, по-видимому, за счет наличия в нем вазоконстриктора. В 5 серии анестезию проводили при помощи лидокаина, предварительно проинкубированного (в течение 12 ч при Т +4°С) с человеческим альбумином (10 мг/мл). У этих животных анестезия не наступала. В 6-й серии животным за 5-7 мин до введения лидокаина вводили 1 мл оливкового масла в полость желудка и 1 мл 1% раствора мочевины внутрибрюшинно. Анестезия у животных не наступала вообще.
Из приведенного экспериментального материала видно, что вне зависимости от введения мочевины, она на продолжительность анестезии, вызванной ли-докаином, практического влияния не оказывает. В экспериментах, где анестезию проводили с помощью лидокаина или скандонеста с параллельным введением в полость желудка 1 мл оливкового масла, получили одновременное обезболивание под действием обоих препаратов, которое прекращалось гораздо раньше на фоне повышения концентрации циркулирующих в крови ЖК по сравнению контролем (1 мл ИР в полость желудка). А введенный животным лидокаин, проинкубированный с альбумином, своего анестезирующего действия не проявлял. Не наступало обезболивание и у животных, которым лидокаин вводили после внутрибрюшинного введения МЧ и внутрижелудочного -оливкового масла. Здесь, по-видимому, срабатывает эффект аддитивности, при котором одно вещество усиливает действие другого.
Из последней серии опытов можно заключить, что МЧ необходимы для элиминации МА, так же, как и ЖК. А механизм элиминации МА можно рассматривать, как действие природных метаболитов, постоянно циркулирующих в нашей крови, и оказывающих детер-гентное действие и хаотропный эффект на связи, образовавшиеся между тканевыми белками и экзогенными соединениями (рис.8).
Вестник интенсивной терапии, 2009 г, №2. Клиническая фармакология
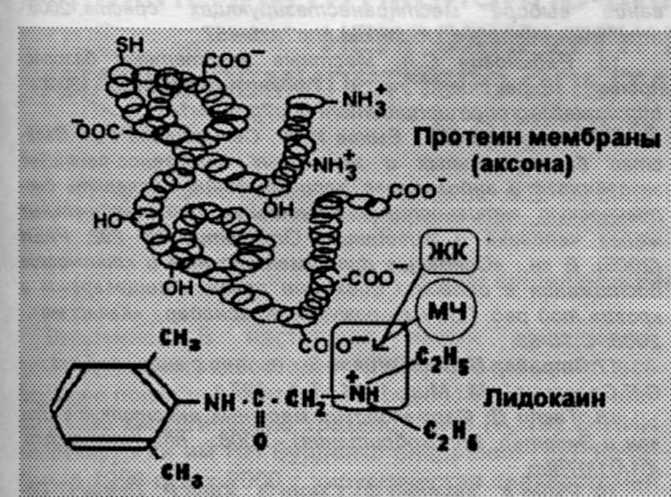
Рисунок 8. Элиминация лидокаина из белково-анвстетического комплекса путем разрыва электростатической связи под действием желчных кислот (ЖК) и мочевины (МЧ).
У врачей, использующих МА для обезболивания различных участков тела, сложилось недостаточно четкое представление о механизме действия амидных местных анестетиков. Оно сформировалось без учета мозаичной структуры плазматических мембран (Singer S.J., Nicolson G.L, 1972), без учета достижений (за последние 2-3 десятилетия) в области гистохимии и биохимии нейрона, фармхимии МА (структуры и химизма их превращений) и неправильного представления их взаимодействия с аксоном.
Так, Malamed S.F. (1997), Бизяев А.Ф. и соавт. (2002), Рабинович OA (2006) и др. авторы считают, что для проявления местноанестезирующей активности препарат должен пройти через фосфолипидную мембрану нервного волокна. А в нем должен произойти гидролиз местного анестетика с освобождением анестетика-основания, действующего непосредственно на ионные насосы. При этом все они придают большое значение константе диссоциации (рКа) анестетика и рН среды, влияющих на проницаемость МА через фосфолипидный слой мембраны. Луцкая И.К. (2002) пишет: «Все анестетики подавляют возникновение и проведение нервных потенциалов за счет обратимой блокады тока ионов Na* внутрь клетки. Блокада натриевых каналов наступает при взаимодействии анестетиков с рецепторами, заложенными в устьях каналов. До того как достичь рецепторов, анестетик проходит липидный слой мембраны. Вот почему анестетик должен растворяться в липидах». Примерно такого же мнения придерживается и Рабинович С.А. (2006а). Однако имеются и более осмысленные концепции, указывающие на то, что МА типа артикаина имеет крайне низкую растворимость в липидах, но хорошо связывается с белками (Rahn R., 1996).
Одной из проблем «проводниковой» анестезиологии является обезболивание при воспалительных процессах. В воспаленных очагах тканей снижается эффективность действия МА, в связи с чем приходится увеличивать их дозировку (Rahn R., 1996; Grigoleit H.-G., 1996; Анисимова Е.Н., 1998; Рабинович С.А., 2006 и др.). Это явление связывают с тем, что в
и др.). Это явление связывают с тем, что в воспаленных тканях рН смещается в кислую сторону, что затрудняет гидролиз местных анестетиков, значительно уменьшая количество анестетика-основания, способного проникать через мембрану нервного волокна. А ухудшение диффузии изменяет способность анестетика вызывать эффективное обезболивание (Yagella J.A., 1991).
Здесь, по-видимому, уместно привести совершенно иное объяснение о складывающейся ситуации в очаге воспаления. В воспаленных тканях из-за развившейся гипоксии нарушается аэробное окисление глюкозы, что приводит к накоплению лактата и пирувата. Они то и, смещая рН, изменяют слабощелочную реакцию среды на кислую. А в кислой среде изменяется соотношение диссоциированных радикалов аминокислот, имеющих аминогруппу (лизина и аргинина) и карбоксильную группу (глутамата и аспартата). При этом уменьшается количество радикалов с отрицательной полярностью, являющихся рецепторами для амидных МА, и увеличивается количество радикалов с положительной полярностью, которые отталкивают эти анестетики с таким же электрическим зарядом. Вот в чем, на наш взгляд, истинная причина снижения активности МА в воспаленном очаге.
Итак, вместо выводов, мы предлагаем при рассмотрении механизма действия местных анестетиков амидного ряда, наряду с общеизвестными фактами и установленными нами механизмами элиминации МА, руководствоваться следующими положениями:
Мембранные интегральные белки выступают из плоскости билипидного слоя, как со стороны внутренней, так и со стороны наружной поверхности плазматической мембраны, а на них имеются функциональные группы преимущественно отрицательной полярности, которые способны вступать во взаимодействие с химическими соединениями противоположной полярности (амидными местными анестетиками).
Амидные местные анестетики (производные лидокаина, артикаина и мепивакаина) заряжены положительно, т.к. в качестве растворителя используется HCI, диссоциирующая на катион Н* и анион СГ. При этом Н* присоединяется к трехвалентному азоту аминогруппы нейтрального анестетика, превращая его в пятивалентный положительно заряженный реагентоспособ-ный анестетик.
Амидные анестетики не растворяются в липидах мембраны аксона, так как её миелинизированные фрагменты покрыты многими слоями шванновской клетки, а под нею нет белков транспортеров для ионов К* и Na+.
Анестезиологическое действие на аксон осуществляется только в местах перехвата Ранвье, где сосредоточены интегральные белки-транспортеры, через которые переносятся различные ионы, простые и более сложные природные органические молекулы. Для МА таких белков нет, а через билипидный слой, не покрытый миелином, беспрепятственно диффундируют Ог, СОг, N2, Н2О, NH3, мочевина и др.
5. Ионы \С и Na+ транспортируются через каналыфермента АТФ-азы, которая одновременно обеспечи-вает и энергией, необходимой для этого активногопроцесса, за счет расщепления АТФ на АДФ и Н3РО4.Существует две теории их транспорта: 1-я - транспорт
К* и Na* осуществляется через один канал в результате антипорта, 2-я - для каждого катиона имеется свой канал.
При инфильтрационной анестезии препаратами местного действия происходит взаимодействие их ионизированных аминогрупп [-NH+-(R)2] с карбоксильными группами (-СОО) аминокислотных радикалов а-субъеди-ниц АТФ-азы, а не с р-субъединицами, так как у последних выступающая часть белка над билипид-ным слоем мембраны прикрыта олигосахаридным компонентом.
После образования электростатической связи между МА и АТФ-азой происходят изменение кон-формации белка фермента, в результате чего перекрываются каналы для потока ионов К* и Na*. Но допускается и второй вариант действия МА, согласно которому он присоединяется к аминокислотному радикалу у входа в канал - прикрывает его устье. Возникает препятствие для транспорта ионов при полной проходимости канала.
Процесс элиминации амидных анестетиков осуществляется естественными метаболитами организма, которые выступают в роли детергента (желчные кислоты) и хаотропа (мочевина). Высвободившиеся МА связываются альбуминами крови, транспортируются в печень, где подвергаются трансформации и затем выведению из организма.
Литература