

Рис.
29.
Суммация мышечных сокращений в ответ
на
2
стимула. Отметка времени
20
мс. Объяснение в тексте.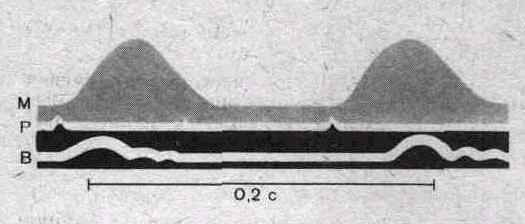
Оптическая регистрация. М — миограмма; Р — oтметка раздражения; В—отметка времени 0,2 с (по Ф. Н. Серкову).
Кривые одиночного сокращения изолированного мышечного волокна сходны по форме с кривыми одиночных сокращений целой мышцы (рис. 28). Отличием является меньшая длительность сокращения одиночного волокна по сравнению с целой мышцей.
Суммация сокращений и тетанус
Если
в эксперименте на отдельное мышечное
волокно или на всю мышцу действуют два
быстро следующих друг за другом сильных
одиночных раздражения, то возникающее
сокращение будет иметь большую амнли
гуду, чем максимальное сокращение
п.р:н, одиночном раздражении. Сократительные
эффекты, вызванные первым и вторым
раздражениями, как бы складываются.
Это явление называется суммацией
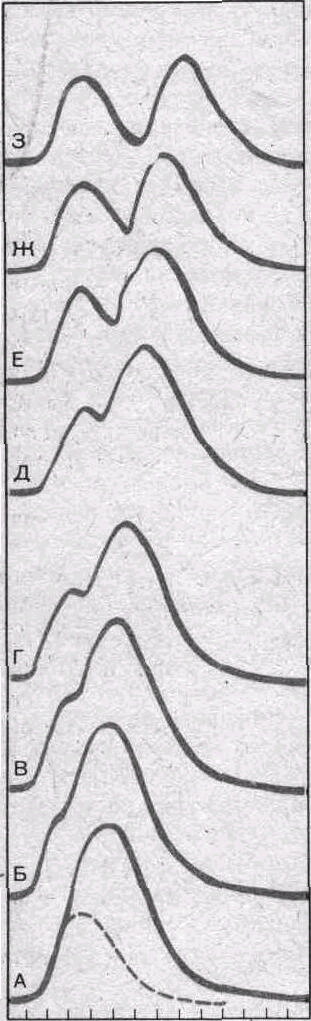
сокращений (рис.
29).
Оно наблюдается как при прямом, так
и при непрямом раздражении мышцы.
Для возникновения суммации необходимо,
чтобы интервал между раздражением имел
определенную длительность: он должен
быть длиннее рефракторного периода,
иначе на второе раздражение не будет
ответа, и короче всей длительности
сократительного ответа, чтобы второе
раздражение подействовало на мышцу
раньше, чем она успеет расслабиться
после первого раздражения. При этом
возможны два варианта. Если второе
раздражение поступает, когда мышца
уже начала расслабляться, то на мио-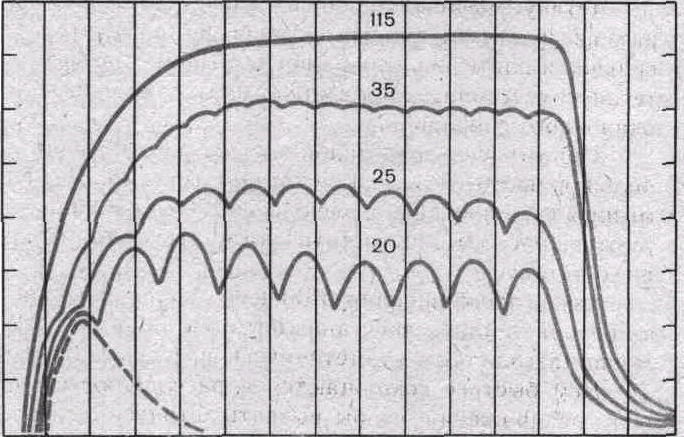
Рис. 30. Сокращения икроножной мышцы при разной частоте раздражения (по Е. F. Жукову).
Одиночное сокращение показано пунктиром. Цифры над кривыми — частоты стимулов. При частотах 20—35 в секунду — зубчатый тетанус, при частоте 115 в секунду— гладкий тетанус.
-Рис.
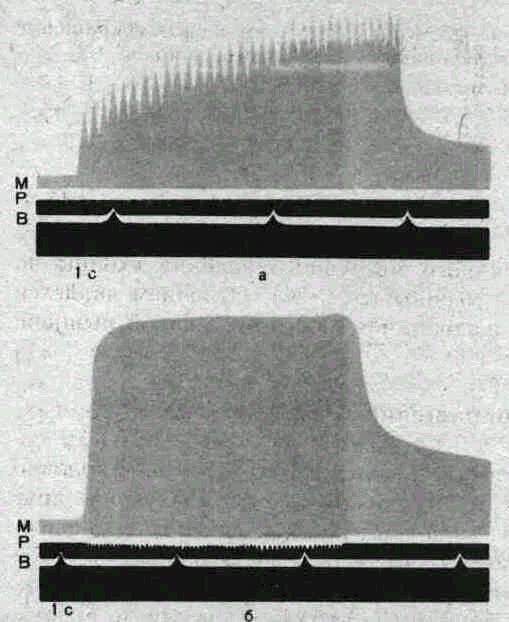
31.
Тетанус изолированного мышечного
волокна (по Ф. Н. Серкову).
а
—
зубчатый тетанус при частоте раздражения
18
Гц; б
—
гладкий тетанус при частоте раздражения
35
Гц; М
—
миограмма; Р
—
отметки раздражения; В
—
отметка времени
1
с.
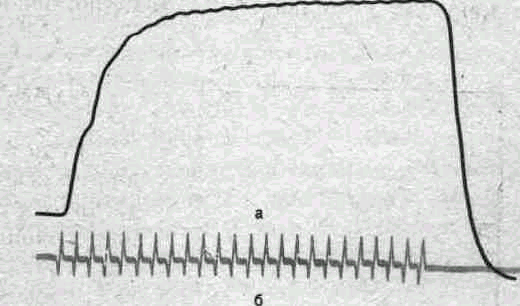
Рис.
32.
Одновременная запись сокращения (а) и
электрической активности (б) скелетной
мышцы кошки при тетаническом раздражении
нерва.
графической кривой вершина этого сокращения будет отделена от вершины первого западением (рис. 29, Ж — Г). Если же второе раздражение действует, когда первое еще не дошло до своей вершины, то второе сокращение полностью сливается С первым, образуя единую суммированную вершину (рис. 29, А—В).
Рассмотрим суммацию в икроножной мышце лягушки. Продолжительность восходящей фазы ее сокращения примерно 0,05 с. Поэтому для воспроизведения на этой мышце первого типа суммации сокращений (неполная суммация) необходимо, чтобы интервал между первым и вторым раздражениями был больше 0,05 с, а для получения второго типа суммации (так называемая полная суммация) — меньше 0,05 с.
Как при полной, так и при неполной суммации сокращений потенциалы действия не суммируются.
Тетанус мышцы. Если на отдельное мышечное волокно или на всю мышцу действуют ритмические раздражения с такой частотой, что их эффекты суммируются, наступает сильное и длительное сокращение мышцы, называемое тетаническим сокращением, или тетанусом. Амплитуда его может быть. в несколько раз больше величины максимального одиночного сокращения.
При относительно малой частоте раздражении наблюдается зубчатый тетанус, при большой частоте — гладкий тетанус (рис. 30, 31). При тетанусе сократительные ответы мышцы суммированы, а электрические ее реакции — потенциалы действия — не суммируются (рис. 32) и их частота соответствует частоте ритмического раздражения, вызвавшего тетанус.
После прекращения тетанического раздражения волокна полностью расслабляются, их исходная длина восстанавливается лишь по истечении некоторого времени. Это явление называется послететанической, или остаточной, контрактурой.
Чем быстрее сокращаются и расслабляются волокна мышцы, тем чаще должны быть раздражения, чтобы вызвать тетанус.