

ГАМК содержится в нейронах спинного и головного мозга. При ее аппликации к различным нейронам ЦНС почти всегда возникает тормозной эффект, вследствие чего ГАМК рассматривают как наиболее распространенный медиатор синаптического торможения. Так, тормозное действие ГАМК было продемонстрировано на клетках коры больших полушарий, нейронах ствола мозга, двигательных нейронах спинного мозга. ГАМК выполняет функцию медиатора при осуществлении как постсинаптического, так и пресинаптического торможения (см. ниже).
Медиаторная функция глицина ограничивается главным образом спинным мозгом, где это вещество выполняет роль медиатора постсинаптического торможения.
Так же как нейтральные аминокислоты, ГАМК и глицин после своего освобождения пресинаптическими окончаниями удаляются из синаптической щели путем захвата нервными и глиальными клетками.
Полипептиды. В последние годы показано, что в синапсах ЦНС медиаторную функцию могут выполнять некоторые полипептиды. К таким полипептидам относятся вещество Р, гипоталамические нейрогормоны, энкефалин и др.
Под названием «вещество Р» подразумевается группа агентов, впервые экстрагированных из кишечника. Эти полипептиды обнаруживаются во многих частях ЦНС. Особенно высока их концентрация в области черного вещества. Наличие вещества Р в задних корешках спинного мозга позволяет предполагать, что оно может служить медиатором в синапсах, образуемых центральными окончаниями аксонов некоторых первичных афферентных нейронов. Действительно, вещество Р оказывает возбуждающее действие на определенные нейроны спинного мозга.
Медиаторная роль других нейропептидов выяснена еще меньше.
Специфические рецепторы мембраны. Для химической передачи в синапсах необходимо существование особых мембранных рецепторов, с которыми реагируют химические медиаторы. Результатом этого взаимодействия является специфическое изменение свойств постсинаптической мембраны, приводящее к возбуждению или торможению постсинаптической клетки.
Роль мембранных рецепторов играют белковые молекулы, обладающие способностью «узнавать» специфические для них вещества и вступать с ними в реакцию. Белковые молекулы подвергаются конформационным изменениям, вследствие чего происходит активация специальных ионных каналов мембраны (ионофоров). В результате этого процесса изменяется ионная проницаемость мембраны, что в свою очередь изменяет мембранную проводимость и приводит к уменьшению или увеличению трансмембранной разности потенциалов—деполяризации или гиперполяризации.
В настоящее время стало очевидным, что рецепторы мембраны довольно быстро обновляются. Они синтезируются, вероятно, в эндоплазматическом ретикулуме, включенном в аппарат Гольджи, и оттуда переносятся к поверхности нервной клетки и включаются в ее мембрану. Весь процесс занимает несколько часов.
Один и тот же медиатор может вступать в реакцию с различными рецепторами постсинаптической мембраны и вызывать противоположные эффекты. Так, в нейронах ЦНС обнаружены мускариновые и никотиновые холинорецепторы, воздействуя на которые ацетилхолин вызывает различные изменения проницаемости постсинаптической мембраны. Показано существование различных рецепторов к катехоламинам. Накапливается все больше данных в пользу существования различных рецепторов к аминокислотам.
Способность одного и того же медиатора вызывать разнонаправленные изменения проницаемости постсинаптической мембраны является причиной того, что одни и те же медиаторы могут или возбуждать, или тормозить различные нервные клетки. В тех случаях, когда влияние химического медиатора более однотипно, как, например, в случае ГАМК и глицина, действие которых почти всегда приводит к увеличению хлорной проницаемости мембраны, функциональный эффект оказывается однозначным (тормозным в случае указанных аминокислот).
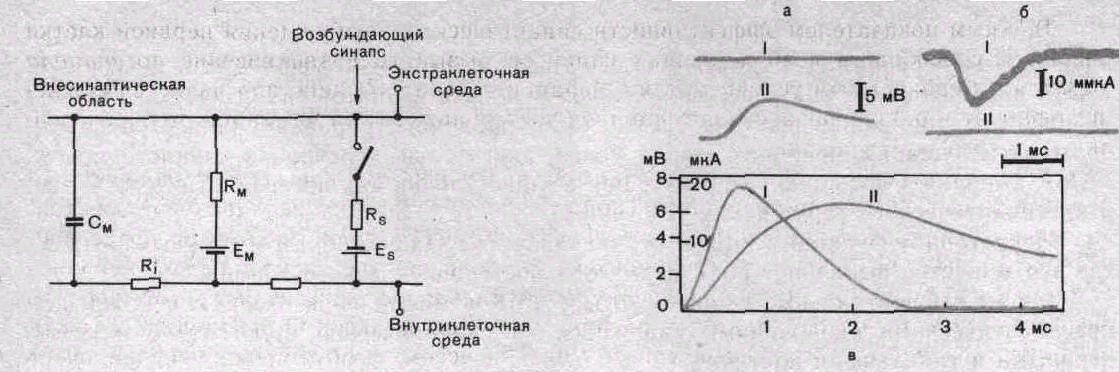
Рис. 66. Эквивалентная электрическая схема возбуждающего синаптического входа.
См—емкость мембраны; Rм, Ri—сопротивление мембраны; Ем—электродвижущая сила мембраны; Es—электродвижущая сила синаптического контакта; Rs—сопротивление синаптического контакта.
Рис. 67. Возбуждающий постсинаптический потенциал (ВПСП) и соответствующий ему постсинаптический ток в мотонейроне спинного мозга кошки.
а: I—ток при отсутствии фиксации потенциала; II — ВПСП; б: 1—ток, И—потенциал во время фиксации; в — график динамики постсинаптического тока (I) и ВПСП (II).
Возбуждение в центральной нервной системе
Хотя один и тот же химический медиатор, действуя на разные рецепторы постсинаптической мембраны в различных нервных клетках может вызывать как возбуждающие, так и тормозные процессы, в ЦНС позвоночных можно выделить синапсы, которые выполняют однозначную функцию—возбуждения (возбуждающие синапсы) (рис. 66). Так, центральные отростки первичных афферентных нейронов всегда оказывают возбуждающее действие на нейроны спинного мозга. Другим примером возбуждающего действия у позвоночных является мотонейрон, активирующий не только мышцы, но и вставочные клетки Реншоу спинного мозга.
В возбуждающих синапсах медиатор, высвобождаемый пресинаптическим окончанием, вызывает развитие локального процесса деполяризации, обозначаемого как возбуждающий постсинаптический потенциал (ВПСП). Указанное название подчеркивает тот факт, что ВПСП развивается в постсинаптической мембране.
В ЦНС млекопитающих ВПСП наиболее подробно изучены в спинальных мотонейронах, где имеется возможность избирательной активации однородных по составу афферентных волокон, которые образуют синапсы непосредственно на мотонейронах. Это позволяет изучать моносинаптические эффекты, не связанные с вовлечением в процесс возбуждения вставочных нейронов (рис. 67).
Амплитуда ВПСП зависит от исходного уровня мембранного потенциала. Смещение мембранного потенциала до величин, близких к нулю, обычно приводит к извращению (реверсии) знака ВПСП, т. е. суммарный постсинаптический ток в этих условиях течет в обратном направлении. Это означает, что активированная возбуждающим медиатором постсинаптическая мембрана становится проницаемой не только для ионов натрия, но и для некоторых других ионов, содержащихся внутри и снаружи клетки. Опыты с введением внутрь нейрона С1~ показали, что при этом амплитуда ВПСП не изменяется. По-видимому, возникновение ВПСП связано с одновременным увеличением проницаемости постсинаптической мембраны для Na+ и К+, а также, возможно, Са2+. Увеличение калиевой проницаемости приводит к уменьшению деполяризации, которая могла бы возникнуть за счет увеличения только натриевой или натриевой и кальциевой проницаемости.
Деполяризация нервной клетки в результате действия возбуждающего медиатора (ВПСП) может быть достигнута не только за счет увеличения проницаемости ее мембраны для Nа+ (или Са2+), но и за счет уменьшения проницаемости для К+.
Важным показателем эффективности синаптического возбуждения нервной клетки является способность возбуждающих синапсов вызывать возникновение потенциала действия. Необходимым условием для генерации потенциала действия является снижение трансмембранной разности потенциалов постсинаптической мембраны до определенного критического уровня.
Условия возникновения потенциала действия в нервной клетке под влиянием синаптического возбуждения в значительной степени обусловлены неодинаковой электрической возбудимостью различных участков мембраны и пространственным распределением различных возбуждающих синапсов. В большинстве центральных нейронов потенциал действия возникает в специальной низкопороговой области (обычно это зона аксонного холмика), откуда он распространяется по аксону и на мембрану соседних участков клетки. Указанный способ синаптического возбуждения нейрона очень важен для его интегративной функции, т. е. способности суммировать влияния, поступающие на нейрон по разным синаптическим путям.