

6 курс / Эндокринология / Инсулиновая_резистентность_и_роль_гормонов_жировой_ткани_в_развитии
.pdfМИНИСТЕРСТВО ЗДРАВООХРАНЕНИЯ И СОЦИАЛЬНОГО РАЗВИТИЯ РФ ГУ ЭНДОКРИНОЛОГИЧЕСКИЙ НАУЧНЫЙ ЦЕНТР
ИНСУЛИНОВАЯ РЕЗИСТЕНТНОСТЬ
ИРОЛЬ ГОРМОНОВ ЖИРОВОЙ ТКАНИ
ВРАЗВИТИИ САХАРНОГО ДИАБЕТА
Пособие для врачей
И.И.Дедов, М.И. Балаболкин, ГГ. Мамаева, Е.М. Клебанова, В.М. Креминская
Москва 2005
Настоящее пособие для врачей посвящено сахарному диабету типа 2, ко торый является медико-социальной проблемой. Расходы на сахарный диабет, включая терапию диабета, его сосудистых осложнений, реабилитация являют ся тяжким бременем бюджета и социального обеспечения общества. Сахарный диабет способствует ранней инвалидизации и высокой летальности, что обус ловлен сосудистыми осложнениями (микро- и макроангиопатии).
Из общего количества больных, на долю пациентов, страдающих сахарным диабетом типа 2 приходится 85-90%. В своей практической деятельности врачи всех специальностей постоянно принимают участие в обследовании и лечении больных, страдающими сахарным диабетом и его различными осложнениями.
Внастоящее время ни у кого не возникает сомнения в том, что в патогенезе сахарного диабета и его осложнений основное значение принадлежит инсулиновой резистентности, которая принимает также участие в патогенезе не только са харного диабета, но и атеросклероза, гипертензии, синдрома склерокистозных яичников и некоторых других заболеваний. Жировая ткань организма является эндокринным органом и многие гормоны, секретируемые ею, непосредственно участвуют в развитии и поддержании инсулиновой резистентности.
Внастоящей работе представлены современные данные литературы и ре зультаты собственных исследований о влиянии различных групп препаратов как на течение сахарного диабета, так и на содержание в сыворотке крови не которых гормонов жировой ткани и на степень выраженности инсулиновой ре зистентности.
Пособие предназначено для эндокринологов, диабетологов, участковых, семейных и врачей общей практики.
Учреждение разработчик.
Эндокринологический научный центр РАМН и кафедра эндокринологии ФППОв ММА им И.М.Сеченова
Составители:
Академик РАН и РАМН профессор И.И.Дедов, профессор М.И.Балаболкин, доктор медицинских наук Г.Г.Мамаева,
кандидаты медицинских наук Е.М.Клебанова и В.М.Креминская
2
ВВЕДЕНИЕ
Инсулинорезистентность - снижение биологического ответа к одному или нескольким эффектам действия инсулина. Однако более часто инсулинорезис тентность определяют как состояние, которое сопровождается сниженным пог лощением глюкозы тканями организма под влиянием инсулина, т.е. это состо яние организма, которое сопровождается резистентностью клеток различных органов и тканей к сахароснижающему действию инсулина.
На чувствительность тканей к инсулину влияют различные факторы, в том числе возраст, наличие избыточной массы тела и особенно распределение жи ровой ткани, состояние артериального давления и наличие гипертензии, нали чие дислипидемии, физическое состояние и тренированность организма, куре ние, ишемическая болезнь сердца и семейный анамнез по диабету.
Общепризнанно, что основная роль в патогенезе сахарного диабета типа 2 принадлежит инсулиновой резистентности и нарушению функции р-клеток. Длительное время не было единой точки зрения относительно преимущест венной значимости одного из указанных факторов в механизмах развития диа бета. Лишь в последние годы благодаря неопровержимым доказательствам большинство исследователей считает, что инсулинорезистентность имеет пер востепенное и основное значение в инициации механизмов, приводящих к раз витию диабета и его сосудистых осложнений.
Таким образом, инсулинорезистентность это состояние, сопровождающееся снижением чувствительности периферических тканей к биологическому дейс твию инсулина и встречающееся не только при сахарном диабете типа 2, но и при других заболеваниях, сопровождающихся нарушениями обмена веществ.
В таблице №1 перечислены состояния, сопровождающиеся инсулиновой
резистентностью. |
|
Таблица 1. Инсулинорезистентность |
|
Физиологическая: |
Эндокринная: |
пубертат |
тиротоксикоз |
беременность |
гипотироз |
ночной сон |
синдром Кушинга |
диета, богатая жиром |
акромегалия |
Метаболическая: |
феохромоцитома |
диабет типа 2 |
Неэндокринная: |
декомпенсация диабета типа 1 |
эссенциальная гипертензия |
диабетический кетоацидоз |
ХПН, цирроз печени |
ожирение |
ревматоидный артрит |
выраженная недостаточность питания |
acanthosis nigricans, раковая кахексия |
гиперурикемия |
сердечная недостаточность |
инсулинвызванная гипогликемия |
миотоническая дистрофия |
избыточный прием алкоголя |
травма, ожоги, сепсис, хирургия |
3
Инсулинорезистентность встречается более чем в 25% у практически здо ровых лиц без ожирения, степень выраженности которой сопоставима с инсулинорезистентностью у больных сахарным диабетом типа 2. Изучая с помощью биостатора распространенность инсулинорезистентности у больных с наруше нием толерантности к глюкозе, сахарным диабетом типа 2, с дислипидемией, гиперурикемией и гипертензией, Е. Вогопа и соавт. (1998) показали, что тако вая встречается при сахарном диабете типа 2 у 83,9% больных; при нарушен ной толерантности к глюкозе - у 65,9%; при гиперхолестеринемии - 53,5%; при гипертриглицеридемии - 84,2%; при снижении липопротеиидов высокой плот ности - 88, 1%; при гиперурикемии - 62,8% и при гипертонии - 58%.
ПАТОГЕНЕЗ САХАРНОГО ДИАБЕТА ТИПА 2
Эссенциальный сахарный диабет типа 2 является гетерогенным и полигенным заболеванием, в патогенезе которого участвуют несколько генетических и внешнесредовых компонентов. Взаимодействие между собой как генетических, так и внешнесредовых факторов является комплексным и не однонаправленным. Гены, определяющие предрасположенность к сахарному диабету типа 2 оперируют уже на самых ранних (эмбриональных) стадиях развития поджелудочной железы, и вовлечены в процессы секреции инсулина и обмена глюкозы в р-клетке, печени и в других тканях организма. Наследование сахарного диабета типа 2 полигенное и в качестве генов-кандидатов рассматриваются следующие гены: ген инсулина, ген рецептора к глюкагону, ген белка, связывающего свободные жирные кислоты, ген гликогенсинтазы, ген белковой фосфатазы типа 1, ген фратаксина, гены глюкозных транспортеров (ГЛЮТ-2 и ГЛЮТ-4), ген р3-адренорецептора, ген гексокиназы типа II, ген фосфатидилинозитол 3-киназы, гены прогормональной сонвертазы и карбоксипептидазы Е, ген амилина, ген рецептора желудочного ингибиторного полипептида, ген островка-1, ген рецептора глюкагон-подобного пептида типа- 1, ген RAD, ген рецептора витамина D, ген белка, связывающего витамин D, ген промотора глюкозо-6-фосфатазы, ген промотора фосфоэнолпируваткарбоксилазы и ген инсулинрезистентного сахарного диабета типа 2, локализованный на длинном плече 20-й хромосомы -локус 20q13.1 -13.2.
Гены, участвующие в предрасположенности к сахарному диабету типа 2, осуществляют это влияние в кооперации с генами, вовлеченными в патогенез ожирения. Если мутация одних генов четко ассоциируется с сахарным диабе том, то мутация других остается как бы «молчащей», не сочетающейся с извес тными нам клиническими и биохимическими признаками заболевания. Более того, мутации генов, сочетающиеся с сахарным диабетом типа 2 неодинаково проявляют себя в различных популяциях, что, видимо, связано с пока неизвес тными нам факторами, которые реализуют имеющуюся мутацию генов в опре деленные нарушения функции отдельных органов и систем, приводя к разви тию определенной клинической картины заболевания.
Несмотря на гетерогенность сахарного диабета типа 2, главными механизма ми в патогенезе заболевания является инсулиновая резистентность и различной степени выраженности недостаточность функции р-клеток. Однако все еще нет
4
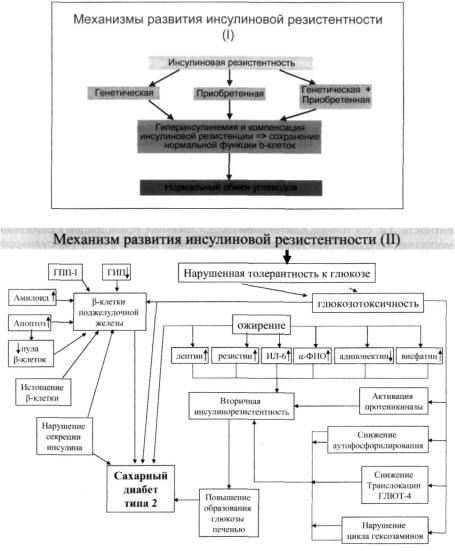
единодушного мнения в том, какой из перечисленных факторов (инсупинорезистентность или дефект функциир-клеток) является первичным. Гипергликемия сама по себе может быть причиной как инсулинорезистентности, так и недостаточнос ти функции р-клеток. Тем не менее, исследования последних лет, в том числе и популяционные, показывают, что инсулинорезистентность является первичной. Механизмы патогенеза сахарного диабета типа 2 представлены на схеме 1.2.
Схема 1. Патогенез сахарного диабета типа 2
5
Причины инсулинорезистентности при сахарном диабете типа 2 гетерогенны. В развитии инсулинорезистентности чётко прослеживается наличие двух ее компонентов: генетического или наследственного и приобретенного. Так, родс твенники первой степени родства с нарушенной и даже с нормальной толеран тностью к глюкозе имеют выраженную инсулинорезистентность по сравнению
слицами контрольной группы. У монозиготных близнецов, имеющих сахарный диабет типа 2 инсулиновая резистентность также более выражена по сравнению
сблизнецами без диабета. Приобретенный компонент инсулинорезистентности проявляется уже в период манифестации диабета. В ряде исследований показа но, что имеющаяся умеренная инсулинорезистентность у родственников первой степени родства при сохранении нормальной толерантности к глюкозе значи тельно усугубляется при нарушении у них углеводного обмена. Аналогичные дан ные получены при проведении исследований у монозиготных близнецов.
Несмотря на то, что инсулиновая резистентность имеет четкую генетическую предрасположенность, до сих пор не идентифицированы точные генетические нарушения, лежащие в основе ее наличия. Это свидетельствует о полигенном ха рактере инсулиновой резистентности. При генетическом обследовании больных, имеющих сахарный диабет, дислипидемию, гипертензию, которые, как известно, являются компонентами синдрома инсулиновой резистентности, было иденти фицировано нескольких точечных мутаций в гене рецептора к лептину, в гене суб страта инсулинового рецептора -1 (СИР-1) и PPAR-y (G. S. Barsh и соавт, 2000).
Механизмы развития инсулиновой резистентности при сахарном диабете типа 2 также гетерогенны. Одной из причин вторичной инсулинорезистентнос ти является глюкозотоксичность или глюкозо-вызванная инсулиновая резис тентность, т.е. состояние длительной гипергликемии, приводящее к снижению биологического действия инсулина. При сахарном диабете типа 1 инсулиноре зистентность развивается как следствие плохого контроля сахарного диабета, а улучшение компенсации углеводного обмена у этих больных сопровождает ся выраженным улучшением чувствительности к инсулину, что требует в свою очередь коррекции суточных доз инсулина, применяемых у больных после до стижения хорошей компенсации диабета. Инсулинорезистентность у больных сахарным диабетом типа 1 в большей степени проявляется в мышечных тканях, носит обратимый характер и имеет прямую корреляцию с содержанием гликогемоглобина в крови. У больных сахарным диабетом типа 2 инсулинорезис тентность обусловлена в большей степени наследственным компонентом, хотя степень ее выраженности также уменьшается при достижении компенсации уг леводного обмена, но в значительно меньшей степени по сравнению с тем, что имеет место у больных сахарным диабетом типа 1.
Инсулинорезистентность сопровождается снижением инсулин-стимулиро- ванного транспорта глюкозы в жировую и мышечную ткани, что было достаточно убедительно показано в исследованиях in vitro (культура фибробластов, преадипоцитов или интактных мышечных клеток). W.T.Garvey и соавт. (1988) первыми показали синергетическое влияние инсулина и глюкозы в механизмах десенситизации глюкозотранспортнои системы в культуре адипоцитов, что позволило им сформулировать новую концепцию развития инсулинорезистентности, что было в последующем подтверждено и другими исследователями. Инкубация
6

мышечных клеток или адипоцитов в среде с высоким содержанием глюкозы (от 10 до 20 ммоль/л), что встречается почти постоянно у больных диабетом, сопровождается прогрессивным снижением глюкозотранспортной системы и развитием инсулиновой резистентности. Это состояние имеет обратимый ха рактер и снижение концентрации глюкозы в инкубационной среде приводит к восстановлению транспорта глюкозы в клетку. Следует указать, что снижению транспорта глюкозы в клетку предшествует короткая фаза, сопровождающаяся нарушением внутриклеточного распределения глюкозы, что позволяет пред положить об имеющихся нарушениях между высоким уровнем доступной для клеток глюкозы (гипергликемия) и дефективным инсулин-чувствительным ме ханизмом распределения глюкозы в клетке.
У больных сахарным диабетом типа 2 синтез гликогена в мышцах снижен на 50% по сравнению с практически здоровыми лицами. Синтез гликогена осущест вляется под контролем гликогенсинтазы, активность которой, в свою очередь, регулируется киназой Зргликогенсинтазы (ген локализуется на хромосоме 19q13.1 -13.2) и киназой Зр-гликогенсинтазы (ген локализуется на 3q13.3-q21.1). Нарушение гена гликогенсинтазы и ее киназ сопровождается инсулиновой ре зистентностью. Синтез гликогена в мышцах представлен на схеме 2
Установлено, что синтез гликогена в мышцах больных СДТ2 снижен на 50% по сравнению с контролем
Ген гликогенсинтазы
Ген киназы 3-а гликогенсинтазы на 19q13.1-13.2 Ген киназы 3-Р гликогенсинтазы на 3q13.3-q21.1
Схема 2. Синтез гликогена в мыщцах
Помимо этого глюкозотоксичность способствует десенситизации р-клеток, что проявляется ухудшением их секреторной активности. Установлено также, что некоторые аминокислоты и, в частности, глютамин значительно влияют на действие инсулина, модулируя поглощение глюкозы. В таких случаях десенситизация глюкозотранспортной системы опосредуется продуктами обмена глюкозаминового шунта. Уже на ранних стадиях этого процесса наблюдается
7

ухудшение транслокации глюкозных транспортеров, главным образом ГЛЮТ-4, в ответ на действие инсулина, а затем и на экспрессию генов глюкозных транс портеров, что, в первую очередь, сопровождается снижением количества мРНК ГЛЮТ-4. Считается, что в процессах десенситизации глюкозной транспортной системы, значительную роль играет незначительный путь утилизации глюкозы в клетке, так называемый гексозаминовый шунт. Глютамин и фермент фруктозо-6 фосфат аминотрансфераза необходимы для конверсии фруктозо-6-фосфата в глюкозамин-6-фосфат и для нормального функционирования этого шунта.
Изучая состояние транспортной системы, осуществляющей поступление глюкозы в клетку, S. Marshall и соавт. (1992) показали, что для десенситизации глюкозной транспортной системы в клетках жировой ткани, инкубированных при высоких концентрациях глюкозы и инсулина, необходим хотя бы в минимальных количествах обмен глюкозы/фосфатов гексоз, т.е наличия функционирования биосинтеза гексозаминов, для инициации которого необходима конверсия фруктозо-6-фосфата в глюкозамин-6-фосфат при участии фермента глютамин: фруктозо-6-фосфат аминотрансфераза или GFAT. При этом было показано, что инсулин осуществляет пермиссивную роль в глюкозо-вызванной десентизации и этот эффект не зависит от комплексирования инсулина с рецептором. Гексо заминовый шунт представлен на схеме 3.
СЖК повышают в митохондриях NADH/NAD+ и ацетилСоА/СоА, что ингибирует ПДГ (пируват дегидрогеназу) с последующим повышением цитрата, который угнетает ФФК (фосфофруктокиназу).
Повышение Г-6-Ф ингибирует активность гексокиназы II (ГК), что приводит к повышению внутриклеточной концентрации глюкозы и снижению ее поглощения мышцей
Схема 3. Взаимоотношения между уровнем глюкозы, СЖК и инсулиновой резистентностью
Поступившая с помощью глюкозных транспортеров (в основном ГЛЮТ-4) во внутрь клетки глюкоза с участием фермента гексокиназы превращается в глю- козо-6-фосфат, который в свою очередь используется: а) для образования гли когена, б) в системе гликолиза (около 95-7%), в) в пентозофосфатном шунте и г) лишь 2-3% - в глюкозаминовом шунте. Таким образом, гексозаминовый шунт
8

функционирует как регулятор десенситизации глкозотранспортной системы в жировых и мышечных клетках, участвуя в механизмах, проявляющихся наруше нием действия инсулина на поглошение глюкозы клеткой при состояниях инсулиновой резистентности. При гипергликемии, вследствии повышенной доступ ности глюкозы во внутрь клетки, увеличивается включение углеродов глюкозы в фруктозо-6-фосфат или уменьшается вхождение глюкозы в систему гликолиза и образования гликогена. Механизм, с помощью которого гипергликемия сни жает скорость поглощения глюкозы в клетку под влиянием инсулина, является гексозаминовым шунтом. Увеличение внутрклеточного количества продуктов гексозаминового шунта (гексозфосфатов) по принципу обратной связи регули рует механизмы поглощения глюкозы в клетку, снижая ее поступление, что и наблюдается при инсулинрезистентных состояниях.
Свободные жирные кислоты (СЖК) оказывают ингибирующее влияние на окисление глюкозы (цикл Рэндла) и участвуют в поддержании и усилении со стояния инсулинорезистентности (схема 4).
Схема 4. Современная гипотеза механизма влияния СЖК на инсулинорезистентность
Однако, как показано, исследованиями последних лет, НЭЖК являются не обходимыми для нормальной глюкозо-стимулированной секреции инсулина. Эффективное и быстрое снижение НЭЖК в крови у здоровых и больных диа бетом снижает базальный уровень секреции инсулина (G. Boden, 1999). Кроме того, снижение уровня НЭЖК в крови у родственников 1 -ой степени родства больных сахарным диабетом типа 2, при длительном приеме аципимокса, со провождалось улучшением секреции инсулина поджелудочной железой и инсу лин-опосредованного поглощения глюкозы периферическими тканями, тогда
9
как инфузия липидов здоровым добровольцам в течение 48 часов приводит к инсулиновой резистентности, хронической гипергликемии, которая сопровож далась гиперинсулинемией. Инсулинотропная активность жирных кислот повы шается со степенью их насыщенности (J.D. McGarry, 1999).
В эксперименте на модели «ожирелых» диабетических крыс (Zucker diabetic fatty rat) показано, что значительное повышение в плазме крови концентрации НЭЖК и триглицеридов в предиабетический период сочетается с резким уве личением содержания триглицеридов в островках поджелудочной железы. Это ингибирующее влияние повышенной концентрации липидов на функцию В-кле ток поджелудочной железы названо липотоксичностью. Интересно, что диета
сограничением жиров, назначенная указанным экспериментальным животным
ввозрасте 6 недель снижает гиперлипидемию, гипертриглицерилемию и на копление липидов в островках поджелудочной железы у этих животных, что со провождается улучшением функции (3-клеток. Липотоксичность или длительное влияние повышенных концентраций НЭЖК на снижение функции В-клеток опос редуется нарушением регуляции индуцируемой NO-синтазы и повышенным образованием N0, а применение различных веществ (лептин или троглитазон), приводящих к снижению содержание триглицеридов в островках поджелудоч ной железы, предохраняет повышение уровня N0 в В-клетках, в ответ на IL-ip и последующие явления цитотоксичности. Не исключается, что липотоксичность может опосредоваться и другими механизмами. Установлено, что при повыше ние аккумуляции жирных кислот в островках поджелудочной железы приводит к ускорению апоптоза в р-клетках и повышению в них синтеза керамидов (М. Shimabukuro и соавт., 1998). В исследованиях in vivo на модели ожирелых диа бетических крыс также показано, что апоптоз принимает участие в развитии недостаточности р-клеток, что проявляется невозможностью коменсировать проявления инсулиновой резистентности (A.Pick и соавт.,1998).
Таким образом, перечисленное свидетельствует в пользу того, что нару шение липидного обмена и избыточное накопление НЭЖК в островках под желудочной железы приводит к снижению их функциональной активности, наблюдаемой при сахарном диабете типа 2. Нарушение пульсирующей сек реции инсулина и влияния инсулина на распределение глюкозы в организ ме также являются факторами способствующими инсулинорезистентности. Механизмы развития инсулиновой резистентности при нарушении жирового обмена (повышение НЭЖК или СЖК) у больных сахарным диабетом типа 2 ,были представлены на схеме 3.
Полученные в последние годы данные позволили внести некоторые кор рективы в гипотезу о механизмах, участвующих в развитии инсулиновой резистентности под влиянием повышенного уровня СЖК в крови, что было представлено на схеме 4.
Как отмечалось выше, биологическое действие инсулина, т.е. влияние на стимуляцию поглощения глюкозы периферическими тканями, опосредуется через инсулиновые рецепторы (схема 5).
10