

4 курс / Общая токсикология (доп.) / Toxikologia_ryb_Lukyanenko
.pdfможно предположить, что в данном случае на первый план выступает фактор индивидуальной устойчивости, определяющий время гибели рыб при неизменной концентрации и времени контакта.
Последующее увеличение температуры на 5°С (с 15 до 20°С) приводит к дальнейшему сокращению времени гибели подопытных карасей (примерно в 3 раза) и заметному увеличению процента гибели. Повышение температуры с 20 до 25°С вызывает 100%-ную гибель карасей и вновь сокращает время гибели почти в три раза.
Таким образом, динамика развития токсического процесса и время гибели рыб при неизменной концентрации яда определяются температурой раствора. Увеличение температуры на 10°С (с 15 до 25°С) в области умеренно высоких и умеренно низких температур (с 5 до 15°С) приводит к более или менее равномерным изменениям как времени, так и процента гибели испытуемых рыб. Это относится и к изменениям латентного периода проявления первой фазы фенольной интоксикации: с увеличением температуры с 5 до 15°С латентный период сократился примерно в два раза, а с 15 до 25°С - в три раза.
Изменение времени гибели рыб при увеличении температуры на 10°С в области нижних (5-15°С) и верхних (15-25°С) температур в растворе фенола 400 мг/л далеко не равнозначно (табл. 26).
Таблица 26
Устойчивость карасей в растворе фенола 400 мг/л при разных температурах
Номер |
Температура, |
Длина |
Масса |
Гибель |
Среднее |
время |
Латентный период |
Достоверные |
|
опыта |
°С |
рыб, см |
рыб, г |
рыб, % |
гибели в |
течении |
двигательного |
различия |
|
|
|
|
|
|
двух суток, ч-мин |
возбуждения, мин |
|
|
|
1 |
5 |
9,8 |
15,1 |
100 |
4-07 |
1,5-2 |
1 от 4 |
и 5 |
|
2 |
10 |
10,4 |
17,2 |
95 |
5-44 |
1,0 |
2 от 4 |
и 5 |
|
3 |
15 |
10,0 |
16,8 |
95 |
5-06 |
1,0 |
3 от 4 |
и 5 |
|
4 |
20 |
9,9 |
15,2 |
100 |
0-56 |
0,5 |
4 от 5, |
3, 2 |
|
5 |
25 |
10,1 |
16,0 |
100 |
0-24 |
0,5 |
5 от 4, |
3, 2 |
|
Примечание. Во всех случаях испытывали по 20 рыб.
Оказалось, что с повышением температуры от 5 до 15°С статистически оправданного изменения времени гибели рыб не произошло, в то время как увеличение температуры с 15 до 25°С привело почти к 13-кратному сокращению времени гибели испытуемых рыб без изменения концентрации яда. В интервале температур 15-20°С изменение на несколько градусов привело к пятикратному сокращению времени гибели, что согласуется с ранними наблюдениями Е. А. Ве-селова (1957). Следовательно, время гибели испытуемых рыб изменяется неодинаково при одном и том же увеличении температуры в области нижних и верхних величин. В области верхних температур ускорение гибели подопытных рыб, обусловленное сдвигом температуры на 10°С, выражено значительно больше, чем в области нижних температур. В противоположность данным, полученным в опытах с пороговой концентрацией фенола 25 мг/л, в настоящей серии опытов латентный период проявления первых симптомов отравления существенно не изменялся во всем интервале изученных температур от 5 до 25°С и кратность различий не превышала двух. Таким образом, характер влияния температуры на устойчивость карасей к концентрациям фенола 25, 100 и 400 мг/л неодинаков. Такие показатели токсикорезистентности, как латентный период интоксикации, скорость и процент гибели рыб, претерпевают различные по выраженности изменения. Графическое изображение связи между температурой и процентом гибели испытуемых рыб при неизменной концентрации яда, представленной на рис. 16, позволяет считать такую зависимость реальной только при концентрации фенола 100 мг/л. Латентный период отравления изменялся с повышением температуры в каждом из растворов фенола, но наиболее выразительные изменения были получены в опытах с пороговой токсической концентрацией яда (25 мг/л). Скорость гибели испытуемых рыб была также различной при изменении температуры от нижних к верхним величинам, однако в растворах фенола 100 мг/л, и особенно 400 мг/л, ускорение гибели рыб было более выражено.
Чтобы понять приведенные здесь данные, освещающие характер и выраженность влияния температуры на устойчивость рыб к ядам, необходимы данные о природе биохимических процессов, развертывающихся в организме при разных температурах, которые, к сожалению, отсутствуют. Приходится ограничиваться лишь общим положением - правилом Вант-Гоффа. Поскольку правило Вант-Гоффа было сформулировано на основе опытов с простыми химическими соединениями, то применительно к сложным биологическим системам количественная сторона этой зависимости претерпевает существенные изменения. Это и было подтверждено результатами опытов. Изменение температуры с 15 до 25°С приводило к сдвигу величины основного обмена не более чем на 25%, в то время как по Вант-Гоффу эти колебания должны достигать 200-300% (Schlieрег, 1950).
61

Рис. 16. Зависимость устойчивости рыб к фенолу от температуры: 4 - концентрация фенола 400 мг/л; В - 100 мг/л; С - 25 мг/л.
Еще более интересны в этом плане ранние наблюдения Н. Уэлса (Wells, 1935) и Щебланова, на основе которых Г. Г. Винберг (1956) считает, что адаптация рыб к низким температурам сопряжена с повышением уровня обмена, а к высоким температурам - с уменьшением его интенсивности. Это утверждение не согласуется с выводами, следующими из правила Вант-Гоффа. Рыбы, по-видимому, имеют некоторые элементы терморегулирующего механизма (Пегель, Реморов, 1957, 1959), благодаря чему они способны замедлять обмен при высоких температурах и ускорять три низких.
Значение предварительной адаптации к той или иной температуре и ее влияние на устойчивость рыб к ядам представляет в этой связи особый интерес. Ориентировочные опыты Букштейга и др. (1955) показали, что время проявления токсического эффекта у окуней, адаптированных к температуре 25°С, при концентрации фенола 20 мг/л равно 20,4 мин, а у неадаптированных оно в два раза меньше - 9,2 мин.
При резком снижении температуры токсического раствора с 15 до 3°С время проявления токсического эффекта концентрации фенола 20 мг/л составляло 4,8 мин, а после 48 ч адаптации к температуре 3°С это время увеличилось в 2,25 раза и было равно 10,8 мин. Время проявления токсического эффекта (концентрация фенола 20.мг/л) у рыб, адаптированных к температуре 25°С, было почти в два раза больше, чем адаптированных к температуре 3°С (20,4 и 10,8 мин, соответственно). Авторы объясняют этот феномен способностью рыб замедлять обмен при высоких температурах и ускорять при низких. Они считают, что при мгновенном повышении температуры с 15 до 25°С общий обмен рыб сильно ускоряется, яд воспринимается быстрее и укорачивается время проявления токсического эффекта. При медленных изменениях температуры раствора токсичность фенола меняется менее значительно.
Продолжая исследования в этом плане, мы учитывали не время проявления токсического эффекта, а время гибели и латентный период отравления. С этой целью карасей адаптировали к температуре 20°С в течение восьми дней, затем их погружали в раствор фенола 100 мг/л с такой же температурой. В контрольных опытах равное количество карасей помещали из воды с температурой 10°С в токсический раствор фенола 100 мг/л с температурой 20°С. Среднее время гибели адаптированных к 20°С рыб оказалось почти в три раза меньше (40 мин), чем у неадаптированных (118 мин) (табл. 27).
Таблица 27
Сравнительная характеристика времени гибели адаптированных к 20°С и неадаптированных рыб в растворе фенола 100 мг/л
Температура, |
Число |
Длина |
Масса |
Гибель |
Среднее время |
Латентный период |
Достоверные |
|
°С |
рыб |
рыб, см |
рыб, г |
рыб, % |
гибели в течении |
двигательного |
различия |
|
|
|
|
|
|
двух суток, ч-мин |
возбуждения, мин |
|
|
5 |
20 |
9,8 |
15,1 |
100 |
4-07 |
1,5-2 |
1 от 4 |
и 5 |
10 |
20 |
10,4 |
17,2 |
95 |
5-44 |
1,0 |
2 от 4 |
и 5 |
15 |
20 |
10,0 |
16,8 |
95 |
5-06 |
1,0 |
3 от 4 |
и 5 |
20 |
20 |
9,9 |
15,2 |
100 |
0-55 |
0,5 |
4 от 5, |
3, 2 |
25 |
20 |
10,1 |
16,0 |
100 |
0-24 |
0,5 |
5 от 4, 3, 2 |
|
|
|
|
|
|
|
|
|
62 |
Вопреки ожиданию, время гибели адаптированных к высокой температуре рыб не только не увеличилось, но значительно уменьшилось по сравнению со временем гибели неадаптированных рыб. Трудно объяснить этот факт на основе представлений, что адаптация к высоким температурам сопровождается уменьшением интенсивности обмена, так как ускорение интоксикации и времени гибели свидетельствуют о возрастании интенсивности обмена. Возможно, что расхождение наших данных с утверждением В. Букштейга и др. (1955) обусловлено различием в реакциях стенотермных и эвритермных рыб на изменения температуры, а также сезонными колебаниями метаболизма, определяющими, по-видимому, выраженность реакции рыб на изменение температуры.
В зимних опытах на карасях не удалось выявить влияния температурного фактора на порог устойчивости рыб и изменение величины минимальной смертельной концентрации яда. Поэтому была поставлена еще одна серия опытов в июне 1964 г. с язем, плотвой и густерой. В качестве токсических агентов использовали фенол, гидрохинон, орто-, метакрезол. Опыты проводились при температуре 7 и 15°С в 200-литровых плексигласовых аквариумах. Определяли величину CL50 и CL100, вызывающие гибель за 24 ч (табл. 28). Изменение температуры токсического раствора в два раза приводило к некоторому хорошо заметному изменению величины обеих концентраций. Выраженность и содержание этих изменений в опытах на разных видах рыб и с различными токсическими агентами оказались неравноценными.
|
|
|
|
|
|
|
|
Таблица 28 |
|
|
|
Сравнительная характеристика величин CL50 |
и CL |
100 при температуре 7 и 15°С |
|
||||
|
|
|
|
|
|
|
|
|
|
|
Вещество |
|
Вид рыбы |
Число рыб |
|
CL50/CL100 при 7°С, |
CL50/CL100 при |
|
|
|
|
|
|
|
|
|
мг/л |
15°С, мг/л |
|
|
Метакрезол |
|
Язь |
92 |
|
8/13 |
11/15 |
|
|
|
» |
|
Густера |
96 |
|
13/9 |
16/14 |
|
|
|
» |
|
Плотва |
48 |
|
20/- |
30/- |
|
|
|
Ортокрезол |
|
Язь |
94 |
|
8/10 |
15/25 |
|
|
|
Фенол |
|
» |
92 |
|
10/15 |
16/18 |
|
|
|
» |
|
Плотва |
90 |
|
25/15 |
40/22 |
|
|
|
Пирогаллол |
|
» |
88 |
|
30/30 |
42/42 |
|
|
|
Гидрохинон |
|
» |
90 |
|
1/- |
|
0,25/0,25 |
|
|
» |
|
Густера |
88 |
|
0,18/0,12 |
0,25/0,22 |
|
|
Вопытах с язем снижение температуры токсического раствора с 15 до 7°С привело к уменьшению
устойчивости рыб и, как следствие этого, снижению величин CL50 и CL100. Устойчивость плотвы к фенолу, а густеры к гидрохинону с понижением температуры увеличилась. Следовательно, уровень устойчивости рыб к токсическому действию различных компонентов сточных вод при одинаковом изменении температуры определяется видовыми особенностями испытуемых рыб, а также сезоном проведения опытов.
Эти опыты показывают необходимость внести существенные коррективы в наши представления о роли температурного фактора в определении степени устойчивости рыб к ядам. Сезонные особенности метаболизма, его цикличность оказывают решающее влияние на токсикорезистентность рыб. Эффект изменения температуры токсического раствора проявляется через существующий в данный момент уровень и направленность метаболизма, складывающиеся в процессе эволюционного развития. Именно поэтому снижение температуры летом и повышение зимой и приводят к одному и тому же эффекту - снижению уровня токсикорезистентности рыб.
Взаключение отметим, что изменение температуры среды обитания рыб является важнейшим экологическим фактором, оказывающим неспецифическое влияние на токсикорезистентность рыб к ядам органической и неорганической природы. Повышение температуры в диапазоне от 5 до 25°С при неизменной концентрации яда в значительной мере изменяет динамику интоксикации, существенно ускоряя ее развитие и исход, а также оказывает заметное влияние на скрытый период отравления. Повышение температуры влияет не только на время гибели, но и на порог устойчивости рыб, снижая величину минимальной летальной концентрации. Следовательно, концентрация яда, губительное действие которого не выявляется при умеренных или низких температурах, может оказаться смертельной при повышении температуры в водоеме. Одновременно с этим следует отметить, что при резком снижении температуры в летне-осеннее время также может проявиться токсическое действие подпороговых концентраций различных компонентов промышленных сточных вод.
Пока трудно установить значение температурного коэффициента Q10 для токсикологических процессов и кратность изменения скорости гибели с повышением температуры на 10°С, поскольку такое повышение температуры в области умеренно низких (от 5 до 15°С) и умеренно высоких (с 15 до 25°С) температур приводит
63
к различному по своей выраженности эффекту. Следует добавить, что изменение температуры воды влечет сдвиги и некоторых других параметров водной среды, в частности меняет уровень кислородного насыщения.
СОДЕРЖАНИЕ КИСЛОРОДА В ВОДЕ
От насыщения воды кислородом зависит нормальная жизнедеятельность рыб и вследствие этого их устойчивость ко многим неблагоприятным факторам внешней среды, включая яды промышленных сточных вод. Уменьшение количества растворенного кислорода в водоеме может быть вызвано разнообразными причинами: бактериальным разложением органических веществ, загрязнением сточными водами предприятий и др. Существенное влияние на уровень насыщения воды кислородом оказывает температура, так как с изменением температуры воды меняется величина растворимости кислорода (табл. 29).
Таблица 29
Растворимость кислорода (в мг/л) в пресной и соленой воде при разных температурах
(по Krogh, 1941)
Температура, °С |
|
Соленость, % |
|
|
|
|
|
|
|
|
0 |
|
10 |
20 |
|
|
|
|
|
0 |
14,7 |
|
13,0 |
11,4 |
10 |
11,5 |
|
10,3 |
9,0 |
15 |
10,3 |
|
9,3 |
8,3 |
20 |
9,4 |
|
8,5 |
7,6 |
30 |
7,8 |
|
7,2 |
6,4 |
При прочих равных условиях растворимость кислорода в пресной воде выше, чем в соленой.
Зимой часто наблюдается резкое снижение содержания кисло рода и его дефицит вызывает массовые заморы, нанося значительные потери рыбному хозяйству. Нередко заморы возникают и летом, главным образом ночью, вследствие усиленного потребления кислорода водной растительностью. Поскольку диффузия кислорода в воде крайне медленна, летние заморы чаще возникают в слабопроточных водоемах.
Потребность рыб в кислороде, необходимом для обеспечения нормального уровня метаболизма, также различна. Есть виды рыб, способные существовать при очень малом содержании кислорода (0,5 мг/л) - карась, сазан, линь. У кумжи, гольяна, форели потребность в кислороде чрезвычайно велика. Пороговая концентрация кислорода для этих видов рыб лежит в области 5 мг/л, а необходимый для жизнедеятельности уровень насыщения воды кислородом- 7-11 мг/л. Внезапное и резкое снижение концентрации растворенного кислорода оказывает губительное действие на многие виды рыб. Тем не менее определение пороговых концентраций растворенного кислорода для разных видов рыб еще не получило должного развития.
Джонс (Jones, 1964) суммировал все разрозненные сообщения по этому вопросу (табл. 30).
Приведенные в табл. 30 данные следует воспринимать лишь как предварительные, поскольку неизвестны условия, при которых они были получены (за исключением температуры).
Установлено, что кроме температуры величина рН, концентрация свободной углекислоты, а также предварительная адаптация к различному содержанию кислорода оказывают влияние на устойчивость рыб к дефициту кислорода и тем самым на величину пороговой концентрации. Особое значение имеет предварительная адаптация рыб к пониженному содержанию кислорода и ее влияние на летальный кислородный предел. А. Дэвисон (по Doudoroff, 1957) выявил способность некоторых лососевых переносить низкие концентрации растворенного кислорода (1,8-2,4 мг/л) после выдерживания их в течение четырех недель в частично обескислороженной воде. Активность рыб при этом несколько снижена, но угнетение может быть снято повышением содержания кислорода до 3 мг/л. М. Шепард (Schepard, 1955) показал, что устойчивость ручьевой форели к дефициту кислорода может быть Повышена путем предварительной адаптации к низким концентрациям кислорода. Кислородный порог при этом снижается почти в два раза. Устойчивость рыбы повышается, а летальный порог снижается после предварительной адаптации к пониженным концентрациям кислорода. Автор предполагает, что этот эффект обусловлен повышением способности рыб поглощать и связывать кислород при низких концентрациях, т. е. речь идет об улучшении транспортной функции крови и повышении сродства гемоглобина крови к кислороду. Это подтверждается устойчивостью рыб к дефициту кислорода и сродством гемоглобина крови к кислороду: чем выше это сродство, тем выше устойчивость к недостатку ч кислорода и ниже кислородный порог. Однако можно дать и другое объяснение этому факту. Возможно, что повышение устойчивости рыб к дефициту кислорода после предварительной адаптации к
64
пониженииному режиму обусловлено снижением уровня активного и стандартного обмена. В пользу этого предположения свидетельствуют результаты опытов А. Г. Минц по адаптации рыб (карп, карась линь, окунь) к пониженному (8-10%) содержанию кислороду в воде. Уровень обмена, показателем которого служила скорость потребления кислорода, повышается после адаптации к высокому кислород, ному режиму и снижается после адаптации к низкому. Выраженность сдвигов в уровне обмена после адаптации к тому или иному кислородному режиму определяется экологическими особенностями испытуемых видов рыб.
Таблица 30
Пороговые концентрации кислорода для рыб
(по Jones, 1964)
Вид |
Концентрация, мг/л |
Температура, °С |
|
|
|
Уклея |
0,68-1,44 |
16 |
Коричневый сомик |
0,3 |
30 |
Кумжа |
1,13 |
6,4 |
» |
1,16 |
9,5-10 |
» |
2,13 |
18 |
» |
2,8 |
24 |
» |
1,28-1,6 |
9,4 |
» |
1,64-2,48 |
17,-2 |
» |
2 9 |
|
Карп |
1,1 |
30 |
» |
0,59-2,5 |
16 |
Кижуч |
1,3 |
16 |
» |
1,4 |
20 |
» |
2,0 |
24 |
Елец |
0,57-1,1 |
16 |
Угорь европейский |
1,0 |
17 |
Серебряный карась |
0,5 |
10 |
То же |
0,6 |
20 |
» |
0,7 |
30 |
Окунь |
1,1-1,3 |
16 |
Радужная форель |
2,4-3,7 |
16 |
То же |
2,5 |
19-20 |
» |
0,83-1,42 |
11,1 |
» |
1,05-2,06 |
18,5 |
Плотва |
0,67-0,69 |
16 |
Лосось |
2,0-2,2 |
8 |
Черный окунь малоротый |
0,63-0,98 |
15,6 |
Голец |
2,0 |
10 |
» |
2,2 |
15 |
» |
2,5 |
20 |
» |
1,52 |
3,5 |
» |
2,4 |
23 |
» |
2,5 |
19-20 |
» |
1,35-2,35 |
15-6 |
Лещ |
2,25 |
20-26 |
Колюшка трехиглая |
0,25-0,50 |
|
Линь |
0,35-0,52 |
16 |
Окунь желтый |
2,25 |
20-26 |
» |
0,37-0,88 |
15,5 |
Значительный интерес представляют данные А. Г. Минц по скорости адаптации пресноводных рыб к измененному газовому режиму. Новый уровень обмена устанавливается спустя 12-18 дней после начала адаптации. В опытах М. Шепарда (1955) показано, что адаптация американской палии к пониженному
65
содержанию кислорода (с 10 до 4 мг/л) наступает уже через 3-12 суток, т. е. даже несколько быстрее, чем в опытах А. Г. Минц.
Большинство исследований по адаптации рыб к низкому содержанию кислорода выполнено на холодолюбивых лососевых (Townsend, Earnest, 1940; Townsend, Cheyne, Davison, 1954; Shepard, 1955).
Устойчивость теплолюбивых пресноводных рыб к низким концентрациям кислорода и значение при этом предварительной адаптации изучены значительно слабее. Можно сказать лишь, что среди пресноводных рыб холодолюбивые менее устойчивы к снижению уровня кислорода, чем теплолюбивые (Job, 1955; Fry, Hart, 1948).
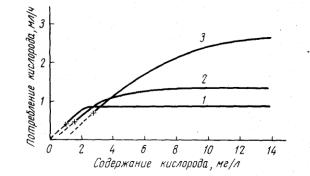
На рис. 17 отражены типичные различия в дыхательной активности холодолюбивых лососевых и теплолюбивых карпов (Fry, 1957).
Рис. 17. Различия зависимости интенсивности дыхания рыб от уровня растворенного кислорода:
1 - Carassius auratus; 2 - Perca flavescens;
3 - Salvelinus fontinalis. Кривые получены
в опытах с 5-граммовыми рыбами при 20° С (по Fry 1957).
Концентрация кислорода 3 мг/л при высоких температурах воды может оказать губительное влияние на малоустойчивые холодолюбивые виды рыб. Резкое снижение концентрации кислорода до 3 мг/л в совокупности с другими неблагоприятными условиями вызывает гибель относительно резистентных видов. Предварительная адаптация к неблагоприятным кислородным условиям приводит к подъему уровня устойчивости и рыбы легко переносят низкие концентрации кислорода (2-3 мг/л) в течение длительного времени. При умеренно низких температурах и других благоприятных условиях концентрации кислорода 1 мг/л и ниже допустимы не только для резистентных видов рыб, но и малоустойчивых, в случае их предварительной адаптации к низкому содержанию кислорода. Характеризуя величину пороговой летальной концентрации кислорода для той или иной группы рыб, необходимо помнить, что она зависит от уровня содержания кислорода в данном водоеме и может заметно отличаться от соответствующей величины для рыб из водоема с другим кислородным режимом. Следовательно, предельно допустимая концентрация кислорода будет различной для рыб разных водоемов и требует специального определения в каждом отдельном случае.
Таким образом, можно считать, что низкое содержание кислорода в воде (0,5-3,0 мг/л) оказывает губительное действие на рыб. Летальная концентрация кислорода зависит от вида рыб и от многих факторов водной среды, из которых особое место занимают яды промышленных сточных вод (Merkens, Downing, 1957).
В свою очередь, дефицит кислорода влияет на интенсивность обмена, приводит к снижению устойчивости рыб ко многим ядам органической и неорганической природы. Выраженность этого влияния определяется степенью связи между интенсивностью обмена у рыб и насыщением воды кислородом. Многочисленными исследованиями, обобщенными в сводке Г. Г. Винберга (1956) и Ф. Фрая (Fry, 1957), установлено, что интенсивность обмена определяется содержанием кислорода в воде. По данным А. Б. Лозинова (1950, 1952, 1953), полученным в опытах на мальках и сеголетках севрюги и осетра, с увеличением содержания кислорода в воде растет скорость его потребления. Но имеется некоторый диапазон концентраций, при которых интенсивность обмена не зависит от уровня растворенного кислорода. Эти факты согласуются с представлениями Т. И. Привольнева (1947) о существовании «кислородной зоны адаптации», т. е. определенного диапазона концентраций кислорода, в пределах которого обмен не зависит от кислородного режима. Рассматривая характер зависимости уровня обмена от содержания кислорода, А. Б. Лозинов предлагает различать «критическую концентрацию» - начальное снижение интенсивности обмена при оптимальной активности и «пороговую концентрацию», вызывающую угнетение основного обмена с последующей гибелью рыб. Сходные соображения были высказаны Ф. Фраем (Fry, 1947). Он подчеркнул, что необходимо дифференцированное изучение зависимости активного и стандартного обмена от степени насыщения воды кислородом. Оказалось, что интенсивность активного обмена более тесно связана с содержанием кислорода в
66

воде, чем основного обмена, и находится в прямой зависимости от уровня кислорода в значительном диапазоне концентраций.
Установлена зависимость интенсивности обмена от совместного действия температуры и содержания кислорода в воде (Fry a. Hart, 1948; Лозинов, 1950; Gibson, Fry, 1954; Job, 1955). С повышением температуры при постоянной концентрации кислорода увеличивается потребление кислорода и интенсивность обмена (рис. 18), т. е. чем выше температура, тем при большем содержании кислорода начинается снижение его потребления. Из данных Д. Фергюсона (по Fry, 1957) следует, что постепенное уменьшение содержания кислорода с 4 до 2 мг/л приводит к снижению интенсивности активного обмена в широком интервале температур от 5 до 25° С (рис. 19). Значительный интерес представляют результаты опытов Д. Грэхем (Graham, 1949), проведенных на сеголетках Salvelinus fontinalls в лаборатории Ф. Фрая. Изучая совместное действие температуры и содержания кислорода на интенсивность обмена, она выявила два интересных факта. Если уровень стандартного обмена постоянно возрастает по мере повышения температуры, то активный обмен увеличивается лишь при подъеме температуры до 20°С, а затем начинает падать, несмотря на последующее повышение температуры. Кроме того, интенсивность метаболизма рыб определяется температурой лишь при оптимальном содержании кислорода в воде. Если насыщение воды кислородом достигает критической величины и падает, то значение температурного фактора в определении уровня обмена полностью нивелируется и на первый план выступает содержание кислорода. Эти факты имеют существенное значение для интер претации результатов многочисленных опытов, направленных на выявление зависимости уровня устойчивости рыб к ядам от содержания кислорода в воде.
Рис. 18. Зависимость активного обмена Amphidon alosoides от температуры и концентрации кислорода:
1-при температуре 5°С; 2- при 10° С; 3-при 15° С (по Fry, 1957).
Рис. 19. Совместное действие концентрации растворенного кислорода и температуры на потребление кислорода Регса flavescens.
Цифрами обозначены концентрации растворенного кислорода:
1 - воздушное насыщение; 2 - 4 мг/л; 3 - 3 мг/л; 4 - 2 мг/л; 5 - контроль (по Fry, 1957).
67

Б. Соулейт, Ф. Пентелов и Р. Базиндель (Southgate, Pentelow, Bassindal, 1933) в серии экспериментов, проведенных на радужной форели с цианидом натрия (0,11 мг/л) и паракрезолом, показали, что в нижних концентрациях кислородного насыщения (от 20 до 60%) устойчивость рыб к ядам быстро падает с уменьшением растворенного кислорода. Если принять в качестве индикатора устойчивости время опрокидывания рыбы, то при 30% кислородного насыщения устойчивость в семь раз ниже, чем при 100% кислородного насыщения. Контрольными опытами авторы показали, что форель при 4,5° С почти трое суток находится без видимого повреждения в воде с 25% кислородного насыщения, что согласуется с более ранним наблюдением Б. Соусгейта (1933). Результаты опытов, представленные на рис. 20 и 21, показывают, что кривые, отражающие связь между устойчивостью форели к обоим ядам и концентрацией кислорода, сходны и вскрывают неспецифический характер этой связи. Авторы приходят к выводу, что если недостаток кислорода сам по себе еще не губителен, то он может существенно снизить устойчивость рыб к различным токсическим компонентам промышленных сточных вод.
Рис. 20. Токсичность цианистого калия для радужной форели при различных концентрациях растворенного кислорода;
t - время опрокидывания рыбы в минутах (слева) и время опрокидывания в минутах (справа)
(no Southgate, Pentelow, Bassindale, 1933).
Рис. 21. Токсичность паракрезола для радужной форели при различных концентрациях растворенного кислорода Условные обозначения те же, что и на рис. 20
(по Southgate, Pentelow, Bassindale, 1933).
Данные Б. Соусгейта и др. (1933) по зависимости устойчивости рыб к ядам от уровня кислородного насыщения были значительно расширены за счет опытов с ядами различных групп органической и неорганической природы (Downing, 1954; Downing, Merkens, 1955; Herbert, Elkins, Mann, Hlemens, 1957; Merkens,
68
Downing, 1957; Weiss, Botts, 1957;' Alderdice, Brett, 1957; Lloid, I960, 1961; Alderdice, 1963 и др.).
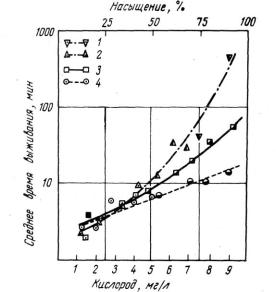
В обстоятельном сообщении К. Даунинг (1954) были изложены результаты опытов по влиянию содержания растворенного кислорода на устойчивость радужной форели к трем различным концентрациям цианистого калия: 0,105; 0,116 и 0,155 мг/л. Всего было поставлено 27 опытов с девятью различными концентрациями кислорода. Было показано, что устойчивость рыб к ядам, показателем которой служит время гибели в растворах цианидов, увеличивается с возрастанием содержания кислорода (рис. 22). Повышение устойчивости рыб более выражено при высоком содержании кислорода и низком уровне токсического агента. Эти факты представляют существенное дополнение к данным Е. Соусгейта и др. (1933).
Рис. 22. Влияние концентрации растворенного кислорода (в мг/л) на период выживания радужной форели в растворе цианистого калия.
Воздушное насыщение дано в %. Для среднего времени выживания дана логарифмическая шкала. Цифрами обозначены подсчитанные величины концентрации вещества в мг/л:
1 - 0,105; 2 - 0,105; 3 - 0,116; 4 - 0,155; (по Downing, 1954).
В совместных работах с Д. Меркенсом (Downing, Merkens, 1955; Merkens, Downing, 1957) К. Даунинг представила новые доказательства зависимости устойчивости рыб к ядам от содержания кислорода в опытах с аммиаком на молоди радужной форели (Salmo gairdneri), окуня (Рекса fluviatilis), плотвы (Rutilus rutilus) и пескаря (Gobio gobio). Концентрация кислорода варьировала от 1,5 до 8,5 мг/л, а аммиака от 0,88 до 8,8 мг/л (N). Время гибели, характеризующее уровень устойчивости рыб, снижалось с увеличением концентрации яда при разном содержании кислорода. В то же время устойчивость рыб возрастала при всех концентрациях яда с повышением содержания кислорода и понижалась с его уменьшением. Авторы отмечают более выраженное влияние на устойчивость рыб низких концентраций кислорода. Указываются видовые особенности изменения устойчивости рыб к ядам в зависимости от содержания кислорода. Так, устойчивость пескаря к аммиаку не претерпевала каких-либо заметных сдвигов в широком интервале концентраций кислорода в сравнении с устойчивостью окуня, плотвы и радужной форели. Подпороговые концентрации аммиака оказали выраженное снижение устойчивости молоди радужной форели к дефициту кислорода, в то время как устойчивость молоди окуня и плотвы осталась без изменений. В отчетах этой же лаборатории (W. P. R., 1958) сообщалось, что в растворах фенолов (от 7 до 14 мг/л) устойчивость рыб снижалась с уменьшением концентрации растворенного кислорода (от 100 до 30% воздушного насыщения). Снижение устойчивости рыб к фенолу оказалось менее выраженным, чем в опытах с цианидами и недиссоциированным аммонием.
Д. Херберт и др. (Herbert et al., 1957) в опытах со сложными сточными водами, содержащими различные комплексы токсических веществ, показали, что снижение концентрации растворенного кислорода с 8 до 4 мг/л приводило к двукратному снижению устойчивости радужной форели (показателем устойчивости служило время гибели рыб в синтетических моющих средствах) (рис. 23). С. Вейс и Д. Ботc (Weis a. Botts, 1957) в опытах с другим токсическим веществом - раствором «Sarin» выявили снижение устойчивости рыб к яду при низком содержании кислорода в воде и увеличение устойчивости по мере его повышения, что согласуется с предыдущими наблюдениями.
69

Рис. 23. Влияние концентрации растворенного кислорода (в мг/л) на период выживания радужной форели (в мин)
в растворе синтетических детергентов (по Herbert ex. al., 1957)
Д. Олдердайсу и Д. Бретту (Alderdice, Brett, 1957) в опытах со сточными водами бумажной фабрики удалось установить изменение пороговой концентрации при изменении кислородного режима (рис. 24). С понижением содержания растворенного кислорода снижалась пороговая токсическая концентрация сточной жидкости с 4,8 до 2,5%. Данные этих авторов удовлетворительно согласуются с данными лабораторного анализа зависимости устойчивости рыб от содержания кислорода в воде.
Рис. 24. Влияние концентрации растворенного кислорода (в мг/л) на токсичность сточных вод бумажной фабрики:
1 - кислород, требуемый для дыхания;
2- имеющийся кислород (по Alderdice а. Brett, 1957).
Изложенные материалы свидетельствуют о существовании тесной связи между содержанием кислорода в воде и уровнем устойчивости рыб к ядам различных химических групп. Снижение уровня кислорода ниже критического порога, будучи само не губительно, сопровождается заметным понижением токсикорезистентности рыб. Каков же механизм реализации этого феномена? В работе французских исследователей Д. Кордье и М. Ворбе (Cordier, Worbe, 19546) показано, что при низком кислородном режиме повышается проницаемость тканей рыб к этиловому спирту. Опыты проводились на плотве при содержании кислорода 1,5-2 мг/л вместо 6 мг/л в контроле, при концентрациях спирта 1,2 и 5 г/л. Об изменении проницаемости судили на основе сопоставления концентрации спирта в теле рыбы и окружающей воде. Испытуемые рыбы, находившиеся в воде с дефицитным содержанием кислорода, содержали больше этилового спирта, чем контрольные. Более высокое содержание этилового спирта у рыб при асфиксии можно объяснить либо повышенной скоростью проникновения яда в организм, либо его замедленной нейтрализацией и выведением, либо тем и другим вместе.
70