

4 курс / Общая токсикология (доп.) / Toxikologia_ryb_Lukyanenko
.pdfРис. 9. Динамика гибели карасей в растворе фенола 50 мг/л.
Работы, выполненные с инсектицидами, также показали, что с увеличением времени испытания изменяются представления о степени токсичности этой группы соединений.
Японские исследователи У. Матсуэ, Т. Эндо и К. Табата (Matsue, Endo, Tabata, 1957) в экспериментах с паратионом на золотых рыбках установили, что при удлинении времени опытов до 30 дней, паратион оказывает токсическое действие при концентрации в 30 раз более низкой, чем 48-часовая летальная концентрация (CL50). На основе полученных данных авторы справедливо высказывают сомнение относительно возможности рекомендовать в качестве допустимой концентрации паратиона для водоемов величину 0,l*CL50, которую в США часто принимают за безвредную.
Испытание токсичности некоторых гербицидов, в частности 2, 4, 5-трихлорфеноксиуксусной кислоты, 4- хлор-2-метилфеноксиуксусной кислоты и других, показало, что продление экспериментов до трех и более недель вызывает токсические явления и даже гибель подопытных групп без существенного изменения концентрации яда
(Bodenstein, Mtiller-Bastgen, 1960).
Всообщении К. Клинглера (Klingler, 1957) приводятся данные, полученные в опытах с нитритом натрия. Хотя вещество малотоксично, однако концентрация 50 мг/л вызывает гибель гольянов в среднем за 14 суток (от 34 ч до 23 суток).
Обстоятельное исследование провел итальянский ученый Р. Маркетти (Marchettii, 1960) на карасях, обработанных различными концентрациями аммиака. Опыты проводились в жесткой воде при температуре 1515,5°С, рН 6,8-7,0. Продолжительность наблюдений - 110 дней. Большинство подопытных рыб погибло в период от 30 до 40 дней. У них были характерные изъязвления, особенно хорошо выраженные на анальном и хвостовом плавниках. Гистологический анализ показал, что это форма типичного некроза. Автор считает, что хотя и нельзя исключить прямого внутреннего действия аммиака, основное действие яда - на защитный слой слизи, вырабатываемой железами, вследствие чего возможно более легкое проникновение бактерийных и грибковых возбудителей в организм.
Попутно отметим, что накожная слизь у рыб играет исключительно важную роль в защите организма от разнообразных повреждающих воздействий.
М. М. Телитченко в опытах с азотнокислым уранилом обнаружил, что основная масса радионуклида аккумулировалась слизью карпа, затрудняя проникновение уранила диффузно через кожу в организм рыбы.
Накопленные экспериментальные данные, полученные в опытах с различными группами химических веществ, убедительно свидетельствуют о ведущей роли фактора времени в определении степени токсичности многих компонентов промышленных вод. Особенное значение длительность опытов приобретает при изучении малотоксичных соединений, вредное действие которых проявляется только после длительного контакта с рыбой.
Вкачестве показателя токсичности того или иного соединения и уровня устойчивости рыб используют время их выживания в токсическом растворе. В ряде работ используют и другие критерии токсичности: 1) время погружения, необходимое для начала токсического процесса; 2) время развития токсического процесса до некоторого хорошо заметного уровня (время опрокидывания); 3) минимальное время экспозиции для наступления токсического процесса (летальное время).
Первый из этих критериев - время погружения - весьма близок по своему содержанию к понятию «латентный период», т. е. промежутку времени между началом контакта рыбы с ядом и первыми признаками отравления. В зависимости от природы испытуемого вещества и вида рыб первоначальные признаки отравления
идлительность латентного периода интоксикации различны. Так, например, у большинства видов рыб первые признаки отравления фенолами: бурная двигательная активность и неориентированное плавание, а у угря и налима - судорожные подергивания мышц туловища. При испытании токсичности растворов тяжелых металлов в качестве первых признаков отравления может быть использовано расстройство дыхания. Время погружения или латентный период весьма различны по абсолютной величине и определяются главным образом механизмами реализации токсического действия. Яды, оказывающие преимущественное действие на центральную нервную систему, имеют короткий латентный период, исчисляемый секундами, а те из них, которые оказывают наружное
41
или локальное воздействие на рыб, имеют более длительный латентный период (минуты, часы и даже дни). Этот критерий может быть использован для характеристики степени чувствительности рыб к яду. Именно поэтому он мало полезен в качестве показателя степени устойчивости рыб к яду или степени токсичности испытуемого вещества.
Второй критерий определяет время наступления одного из наиболее показательных признаков отравления. Чаще всего в качестве такого признака используют потерю рефлекса равновесия, т. е. «время опрокидывания», именуемое также «временем проявления» (manifestation time). Согласно К. Вурману и X. Вокеру (1948) под временем проявления имеется в виду время, в течение которого рыба теряет способность движения и может долго удерживать равновесие в проточной воде. Широкое использование этого критерия для оценки токсичности вещества встречает затруднения. Прежде всего, в случае испытания токсичности веществ, не влияющих на нервную систему (большинство солей тяжелых металлов, щелочи, неорганические кислоты), потеря рефлекса равновесия наступает обычно незадолго до гибели. Кроме того, при действии на рыб других веществ, например цианидов, в качестве наиболее характерных признаков отравления может быть использовано изменение интенсивности потребления кислорода.
Даже имея дело с ядами, вызывающими потерю рефлекса равновесия, едва ли имеет смысл использовать в качестве критерия токсичности время проявления. Опыты показывают, что при изучении токсичности ядов фенольного ряда для различных видов рыб в растворах от 10 до 50 мг/л (в зависимости от вида) наблюдается волнообразное течение интоксикации, при котором периоды потери рефлекса равновесия и полного обездвижения чередуются с появлением кратковременной способности к импульсивным перемещениям в боковом положении. Эти наблюдения в первую очередь относятся к резистентным видам рыб, среди которых следует назвать карася. Но даже в краткосрочных опытах (Лукьяненко и Флеров, 1965) по острому отравлению малоустойчивых видов рыб (форель, окунь) время проявления весьма вариабильно, что не дает возможности с достоверностью судить о степени устойчивости рыб или степени токсичности вещества. Этот критерий, ограничивая длительность наблюдений несколькими часами, не позволяет уловить качественные и количественные особенности протекания токсического процесса при изменении концентрации ядовитого вещества.
Имеются и другие серьезные препятствия для использования этого критерия. Во-первых, нет никаких сведений относительно характера связи между временем проявления и временем гибели рыб результаты наших опытов, полученные на большом количестве испытуемых рыб, показывают, что при значительном, иногда десятикратном увеличении или уменьшении концентрации яда в растворе время проявления претерпевает незначительное редко превышающее двукратное изменение. При этом, соотношение между временем проявления и времени гибели испытуемых рыб оказывается различным как при разных концентрациях токсического вещества, так и при разных веществах даже в опытах на одном виде рыб. Во-вторых, при равном времени проявления время гибели весьма заметно колеблется для разных видов рыб. Поэтому можно считать, что в практическом отношении использование критерия время проявления при оценке токсичности вещества мало оправдано.
Третий критерий - «летальное время» определяется на основе измерения времени экспозиции, приводящего к развитию полностью или частично необратимого отравления. Этот критерий токсичности важен в тех случаях, когда перед исследователем стоит необходимость изучения спорадических загрязнений водоема. Вместе с тем токсикологическое содержание критерия летальное время не вполне ясно. В статье Вурмана и Вокера, посвященной изучению токсичности аммония и цианидов, авторы делят период погружения рыбы в токсический раствор (до смерти) на два этапа: первый этап (латентный) включает в себя период нахождения рыбы в токсическом растворе до потери обратимости; второй этап (летальный) характеризуется периодом необратимого отравления. Такое деление представляет определенный интерес в плане оценки обратимости токсического процесса, однако оно не позволяет характеризовать качественный состав симптомокомплекса отравления, фазность его течения и не дает оснований для суждений о степени резистентности испытуемых рыб. Неудачно обозначение первого этапа «латентным», так как обратимость токсического процесса возможна при весьма глубоких и явных признаках отравления, которые нельзя назвать скрытыми. По-видимому, этот вопрос нуждается в дальнейшей экспериментальной доработке и подборе соответствующей терминологии.
Четвертый критерий - «время выживания» (survival time) представляет собой время, необходимое для развития смертельного отравления. Нам представляется, что время гибели является одним из наиболее полноценных критериев токсичности различных веществ и динамики изменения токсичности в зависимости от концентрации вещества в опытах с острым отравлением.
Столь дифференцированный учет всех этапов развития токсического процесса во времени позволяет не только наиболее полно и точно характеризовать границы токсичности того или иного компонента промышленных сточных вод для рыб, но и точно измерить латентный период, скорость развития и время гибели испытуемых рыб. В современных работах по токсикологии рыб используют только один или два из рассмотренных критериев токсичности вещества, что весьма затрудняет сопоставление данных, полученных в разных лабораториях.
Успешная разработка многих вопросов токсикологии рыб, связанных с определением степени токсичности различных компонентов промышленных сточных вод, требует унификации используемых критериев для
42

сопоставимости литературных сведений и координации научно-исследовательских работ по этой тематике.
Рис. 10. Динамика гибели карасей в растворе фенола 100 мг/л.
Определенный интерес в этом плане может представить «среднее время гибели», подобно тому, как при изучении фармакологической активности соединений используется «среднее эффективное время». Время гибели при неизменной концентрации яда дает широко рассеянный вариационный ряд. Распределение частот в таком ряду хотя и приближается к нормальному, однако нередко существенно отклоняется от него.
Если по оси ординат откладывать процент гибели рыб, а по оси абсцисс промежутки времени, в течение которого наступает летальный эффект, то получается кривая зависимости между временем и процентом погибших животных, которая получила название кривой «время-действие» (time-action). Часто она имеет S- образную форму (как и кривая «концентрация-действие»), подобную той, которую мы получили в опытах с фенолом на карасях (рис. 10). Нередко эти кривые бывают и другой формы (рис. 11).
Рис. 11. Динамика гибели карасей в растворе фенола 200 мг/л
Способы выпрямления S-образной кривой уже были нами изложены, один из них основан на том, что по оси абсцисс вместо времени откладывают его логарифмы, а по оси ординат вместо процента погибших животных - соответствующие пробиты.
Иногда зависимость время - действие имеет форму прямой " (рис. 12). "
Рис. 12. Динамика гибели сегодлеток карася в растворе фенола 50 мг/л
43

Имеется несколько методов (Bliss, 1935; Litchfild, 1949), котоые могут быть применены для исследования выпрямленных прямых и определения среднего летального времени (LT50), а также стандартной ошибки этого показателя. Обычно на графике находят точки, соответствующие LT16, LT50 и LT84 (т. е. время гибели 16, 50 и 84% испытуемых животных). Величину стандартной ошибки в этом случае определяют либо по графику (функция наклона прямой), либо по формуле:
ХАРАКТЕР ЗАВИСИМОСТИ «КОНЦЕНТРАЦИЯ-ВРЕМЯ» И НЕКОТОРЫЕ КОЛИЧЕСТВЕННЫЕ ВОПРОСЫ ТОКСИКОЛОГИИ РЫБ
Зависимость «концентрации-время» (concentration-action) приобрела в токсикологии рыб особое значение. Еще в 1917 г., обстоятельно изучив характер зависимости между концентрацией вещества (соли, кислоты,
наркотики) и временем выживания, Е. Пауэре (Powers) в опытах на золотой рыбке установил, что выше определенной области концентраций время выживания испытуемых рыб обратно концентрации и предложил формулу:
(С – a) t = K
где |
С – концентрация; |
|
а - теоретический порог; |
|
t - время выживания; |
|
К – постоянная |
При нижних концентрациях, однако, измеряемое время выживания короче, чем это следует из уравнения, а при высоких больше, чем теоретическая величина. Если составить график, в котором концентрация откладывается по оси абсцисс, а время выживания по оси ординат, то при интерполяции экспериментальных данных кривая приобретает более или менее сигмоидную форму и лишь средняя ее часть будет прямой (рис. 13). Если продолжить эту прямую до пересечения с осью абсцисс, то точку пересечения условно можно назвать «теоретическим порогом токсичности».
Рис. 13. Отношение между времением выживания и концентрацией: а – теоретический порог токсичности (по Powers, 1917)
Подтверждение выводов Е. Пауэрса было дано в работах Александера, Б. Соусгейта и Р. Базинделя (Alexander, Southgate, Bassindale, 1935) и в более поздних сообщениях К. Вурмана и X. Вокера (1950), Д. Херберта и Д. Меркенса (1952). Александер и др. (1935) на основании опытов с цианистым калием, проведенных на радужной форели, построили график, который сходен с графиком Пауэрса. В работе К. Вурмана и X. Вокера (1950) описываются результаты опытов, цель которых установить зависимость между концентрацией фенола и временем гибели различных видов рыб (форель, окунь, гольян). При количественной оценке полученных данных авторы в качестве исходной использовали формулу:
Cnt = K,
44

где |
С - концентрация яда; |
|
t - время гибели; |
|
n и К. - константы. |
Полученные экспериментальные данные не совпадали с расчетными, вычисленными по этой формуле, поскольку фенол (равно как и другие яды) имеет пороговую концентрацию, ниже которой токсический эффект не проявляется, и минимальное время отравления даже при больших концентрациях. В связи с этим функция Сnt=K представляющая гиперболу с равноугольными асимптотами, претерпевала некоторые изменения, если учитывалась Cs (пороговая концентрация) и минимальное время токсического эффекта - Ts. С учетом этих поправок формула приобретала следующий вид:
(Тm – Ts) (C – Cs)n = K.
При логарифмировании и перестановке получаем
lg (Tm – Ts) = K – n lg(C – Cs)
Иными словами, авторы получили уравнение прямой. График, построенный путем нанесения логарифмов отдельных токсических концентраций (уменьшенных на пороговую концентрацию) и логарифмов времени проявления (уменьшенных на минимальное время реакции), соответствует графику, в основу которого положены экспериментально найденные величины.
В сообщении Д. Херберт.а и Д. Меркенса (1952) по токсичности цианидов для ручьевой форели указывалось, что получена сигмоидная кривая, аналогичная описанной Е. Па-уэрсом. Только вместо обратной величины индивидуальных периодов выживания применялась обратная величина среднего периода выживания. Авторы сопоставили графики, полученные при нанесении логарифмов времени выживания и обратных величин времени выживания, и пришли к выводу, что логарифмическое преобразование имеет явное преимущество. В своей работе Д- Херберт и Д. Меркенс приводят график (рис. 14), где средние периоды выживания при различных концентрациях цианида (в мг/л) нанесены в логарифмическом масштабе. Для определения среднего времени выживания они использовали не обычную статистическую среднюю, а среднее геометрическое из периодов выживания при каждой концентрации яда, найденное по формуле
|
M. S. T. = antilog (lg T / N) , |
где |
М. S. T. (medial survival time) – среднее время выживания; |
|
Т – время выживания каждой рыбы; |
|
N - число испытуемых рыб |
Рис 14. Отношение между концентрацией цианидов и временем выживания радужной форели при температуре 17,5°С
По оси абсцисс – концентрация CN, мг/л;
45
по оси ординат – время опрокидывания (логорифмическая шкала) (по Herbert, Merkens, 1952)
В этом случае прямолинейная часть кривой, как и в опытах К. Вурмана и Х. Вокера (1950), могла быть
представлена уравнением
Сnt=K
В приведенных работах авторы имели дело с каким-то одним ядом, химическая природа которого известна. Гораждо сложнее рассчитывать токсичность различных комбинаций ядов, встречающихся в сточных водах. Д. Олдердайс и Д. Бретт (Alderdice, Brett, 1957) опубликовали результаты своих наблюдений по токсичности отходов целлюлозного производства для молоди лосося и вывели специальный гиперболический эквивалент для прямой линии:
U = a + βV,
где U – обратная величина средних периодов выживания;
V – обратная величиная концентрации сточного материала.
Асимптота β/а дает оценку концентрации сточного материала, безопасную при различных продолжительностях воздействия. Д. Херберт и др. (Herbert a. Al., 1957) изучал токсичность домашних моющих средств для радужной форели. При нанесении на график логарифмов концентрации моющих средств против логарифмов средних периодов выживания была получена прямая линия, выраженная уравнением:
|
lg T = K – n lg C, |
где |
Т – геометрическая средняя периода выживания; |
|
С – концентрация активного материала, мг/л |
|
К = 4,93; |
|
n = 3,71. |
Хотя S-образность кривых «концепция – время» встречается довольно часто, тем не менее экспереминтальные данные не всегда интерполируются с кривой этого типа. Имеется ряд веществ (кофеин, пиридин, нитрит натрияб хлорид магния и т.д.), для которых интерполяция эксперементальных данных с сигмаидной кривой мало оправдана, особенно в области токсичности высоких концентраций (на изгибе сигмоиды). К. Карпентер (Carpenter, 1927) выявил отличие кривой, характеризующей форму зависимости времени гибели испытуемых рыб от концентрации ряда тяжелых металлов, при сопоставлении её с кривой Пауэрса. В этих опятах время выживания (t) было свзано с концентрацией (С) уравнением:
(1/t) lg (1/C) = K,
где К – постоянная.
Д. Джонс (Jones, 1939, 1957) показал, что и для солей серебра, птути, кадмия, никеля, кобальта, соляной кислоты кривые, иллюстрирующие зависимость «концентрация-время» мало похожи на сигмоидные кривые. Г. Гриндлей (Grindley, 1946) обнаружил, что отношение «концентрация-время опрокидывания» для гольяна, помещенного в раствор мышьяковистого натрия, динитрофената натрия или пикрита натрия, было линейным только в определенной области высокотоксичных концентраций. Сходные результаты были отмечены и в опытах на радужной форели при испытании токсичности хромата и бихромата калия.
Изучая характер зависимости между концентрацией фенола и временем гибели карасей, мы получили кривую, существенно отличающуюся от S-ооразной кривой Пауэрса (рис. 15). Можно считать, что форма кривых «концентрация - время» весьма вариабильна для различных ядов и в известной мере зависит от механизма их действия на организм. Время выживания рыб в токсических растворах, вероятно, определяется механизмом действия ядов и большим разнообразием других факторов. Поэтому следует с известной осторожностью оценивать первые попытки определять токсичность неиспытанных концентраций исходя из предположения о математическом характере связи «концентрация-время». Графический метод оценки токсичности неизученных концентраций, предложенный Пауэрсом, путем продолжения прямой части S-образной кривой до абсциссы, на которой нанесены концентрации яда, также встречает ряд затруднений. Опытами Д. Джонса (1939) показана опасность определения поооговых коннентраций только на прямолинейной части кривой и экстраполяцией этих данных за пределы, в которых имеет место прямолинейное отношение. Он обнаружил, что выше области концентраций 0,04-0.0015N токсичность раствора азотнокислой меди [Сu(NО3)2] для головастика жабы
46

возрастала с разведением. При нижних концентрациях время выживания удлинялось и было одинаковым при 0,0003 и 0.04N. К сожалению, автор не указывает статистическую проверку достоверности опытных данных.
Рис. 15. Отношение между временем выживания и концентрацией яда. По оси абсцисс - концентрация яда (в мг/л), логарифмическая шкала; по оси ординат - среднее время выживания рыб, ч;
1- теоретически ожидаемая кривая; 2 - опытная кривая.
В наших опытах по определению токсичности фенола для рыб установлено, что в области концентраций 800-200 мг/л время гибели возрастало с разведением, но при дальнейшем разведении (200100 мг/л) токсичность резко возрасла и в растворе фенола 100 мг/л караси жили значительно меньше времени, чем в растворе 200 мг/л (см. рис. 15). Статистическая обработка результатов по методу Вилкоксона выявила достоверность обнаруженных различий в токсичности сопоставляемых концентраций фенола. В последующем это наблюдение подтвердилось и в опытах с другими видами рыб, более низкие концентрации яда обладали более высокой токсичностью (судя по времени выживания). Следовательно, время выживания испытуемых рыб не всегда возрастает с увеличением разведения. Из наших данных следует также, что скорость снижения токсичности при разведении изменяется весьма неравномерно.
Изучая зависимость токсического эффекта от концентрации яда и времени его действия, Е. А. Веселов (1965) пришел к выводу, что для расчета токсичности неиспытанных концентраций (по времени гибели) может быть использована формула Габера:
W = Ct,
где |
W - токсический эффект; |
|
С - концентрация яда; |
|
t - время действия яда. |
Величина произведения Ct получила название «токсического коэффициента».
Е. А. Веселов считает, что эта формула в ряде случаев применима при обработке данных, полученных на рыбах, но делает оговорку, что произведение Ct сохраняет относительное постоянство только в одной зоне концентраций (летальная или сублетальная). В пределах одной зоны, в случае известной величины W,
W можно рассчитать время гибели t = W / С для веществ с быстрым токсическим эффектом, в качестве которых автор указывает на сернокислую медь и фенол.
Формула Габера, широко применявшаяся ранее, в последние годы редко используется, поскольку даже в пределах одной концентрации в опытах с такими веществами, как гексан, толуол, бензол и др., характеризующимися замедленным токсическим эффектом, величина W значительно колеблется. Использование формулы Габера для расчета токсичности неиспытанных концентраций только в какой-либо одной узкой зоне и притом в случае, если вещество характеризуется быстрым токсическим действием, в значительной мере обесценивает ее практическую значимость. В лаборатории В. М. Карасика (1944) в опытах на мышах было установлено, что при одном и том же времени воздействия фосгена, концентрации, вызывающие гибель отдельных животных, различаются в 4 раза, а в опытах с дифосгеном на кошках амплитуда этих колебаний оказалась равной 6,81. Поэтому В. М. Карасик, вслед за Н. В. Лазаревым (1938), ставит под сомнение возможность широкого использования формулы Габера для количественной интерпретации отношений между
47

концентрацией яда, временем его действия и токсическим эффектом.
В нашей лаборатории, в опытах на различных видах рыб, показано, что фактор индивидуальной устойчивости играет еще большую роль в определении величины W, чем у теплокровных. Даже в пределах одной зоны летальных концентраций фенола произведение Ct под влиянием индвидуальных колебаний устойчивости рыб к яду может меняться в 5-9 раз, поэтому едва ли формула Габера может быть достаточно эффективна «для более точного определения предельно допустимых концентраций ядовитых компонентов промышленных сточных вод».
Установление предельно допустимых и особенно пороговых концентраций промышленных сточных вод - задача весьма сложная. Трудно предполагать, что время выживания всегда будет увеличиваться при разведении, а токсичность - уменьшаться. Необходима дальнейшая экспериментальная разработка этого важнейшего вопроса.
При рассмотрении количественных вопросов токсикологии рыб необходимо обратить внимание на математическую обработку экспериментальных данных, чтобы обеспечить объективную оценку и удобство сопоставления многочисленной информации. Применение статистических методов оценки полученных данных в токсикологии рыб все еще не нашло должного распространения. В настоящее время имеются единичные сообщения (Grindley, 1946; Wuhrmann, Woker, 1950; Trama, 1956), в которых авторы, характеризуя токсичность вещества или степень устойчивости рыб, используют, помимо средней величины, некоторые другие параметры вариационного ряда.
Между тем сама по себе средняя арифметическая имеет ограниченную ценность, поскольку не дает представления о вариабиль-ности учитываемого признака. Кроме того, она в большой мере зависит от особенностей распределения на концах ряда, где частоты бывают наименее надежны и точны. В связи с этим нередко, наряду с определением средней арфиметической
М = Σх / n,
где Σх - сумма отдельных вариант; n - число наблюдений, указывают также центральную варианту ряда - медиану (Me) и наиболее частую варианту - моду (Мо). Сопоставление медианы и моды со средней арифметической позволяет также составить представление о степени симметрии распределения членов данного вариационного ряда - в случае совпадения этих трех величин имеют дело с нормальным распределением.
Поскольку главной особенностью любой выборочной совокупности является наличие вариации между ее членами, возникает необходимость измерения степени этой изменчивости. Нередко с этой целью указывают амплитуду ряда распределения - разность между наименьшей и наибольшей величинами, получивших название лимитов ряда. Сопоставление лимитов ряда не полностью характеризует степень изменчивости внутри вариационного ряда. Поэтому для определения характера и степени вариации или рассеивания используют среднее квадратическое (стандартное отклонение)
и дисперсию
где Σх2 - сумма квадратов отдельных измерений; n - число наблюдений;
М - средняя арифметическая.
Строго говоря, среднеквадратическое отклонение (δ) характеризует амплитуду колебаний средней арифметической, представляя абсолютную величину от средней арифметической. При сопоставлении степени вариации отдельных рядов распределения времени гибели (или другого показателя), измеренных в разных единицах, удобнее использовать коэффициент вариации (υ), показывающий, какую долю средней арифметической составляет среднеквадратическое отклонение. Коэффициент вариации рассчитывается по формуле:
Основные показатели изменчивости величин, составляющих ряд распределения (δ и υ) такие же важные параметры, как и средние величины. Без использования этих показателей,- оперируя лишь средними, можно
48

сделать ошибочное заключение о степени токсичности двух исследуемых веществ.
Таким образом, для описания ряда распределения полученных данных необходимо учитывать не только среднюю арифметическую, медиану и моду, но и распределение частот и их вариабильность, оцениваемую средним квадратическим отклонением и коэффициентом вариации.
Обычно исследователь имеет в своем распоряжении экспериментальные данные, полученные на ограниченном количестве испытуемых рыб, представляющие случайные малые выборки, относящиеся к обширному ряду, именуемому генеральной совокупностью. Поэтому необходима экстраполяция результатов, полученных в выборочной совокупности. Опыты показывают, что точность выборочной средней определяется объемом выборки и степенью вариабильности регистрируемой величины. Однако точность выборочной средней изменяется прямо пропорционально не объему выборки, а квадратному корню из этого объема, т. е. если объем выборки увеличился в 100 раз, точность средней возрастает в 10 раз. Это обстоятельство следует иметь в виду при определении числа испытуемых рыб в токсикологических опытах для того, чтобы выявить эффективную концентрацию яда и среднее время токсического эффекта (Вурман и Вокер, 1950).
Критерием точности средней арифметической служит средняя ошибка (mм) средней арифметической представляющая частное от деления среднего квадратического отклонения от выборочной средней на квадратный корень из числа наблюдений в выборке.
Один из наиболее важных моментов при статистической обработке экспериментальных данных - определение достоверности различия между сравниваемыми величинами. Оно необходимо при характеристике относительной токсичности различных компонентов промышленных сточных вод, а также при оценке роли факторов водной среды в изменении этой токсичности. Оценка достоверности выявленных различий имеет первостепенное значение при описании экологических, видовых и возрастных особенностей устойчивости рыб к ядам.
Существует несколько способов определения достоверности различия. Согласно одному из них различие считается достоверным, если
Другой спосов основан на использовании так называемого критерия t.
где |
М1 и М2 – сравнимые средние арифметические; |
|
- суммы квадратов отклонений членов вариации ряда от их средних |
арифметических;
n1 и n2 – количество наблюдений.
Полученную велицину t сопоставляют со значением t, которое указано в строке с определенным количеством животных, вычисленном по формуле
F = n1 + n2 - 2
по специальной таблице (Weber, 1957)
Если число испытуемых животных в двух сопоставляемых сериях опытов одинаково, т.е. n1 = n2, величина t может быть установлена по более простой формуле:
В некоторых случаях (Wuhrmann, Woker, 1950; Herbert, Mer-kens, 1952) опытным путем удавалось показать, что распределение частот времени проявления или времени опрокидывания соответствует нормальной кривой, однако чаще в работах по токсикологии рыб нормальное распределение лишь предполагается без
49
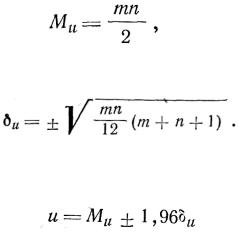
достаточных экспериментальных данных. Поэтому в тех случаях, когда характер распределения полученных экспериментальных данных неизвестен, предпочтительнее использовать в качестве показателя достоверности различия двух рядов распределения непарамётрйчёский критерий Вилкоксона, основанный на числе инверсий. Для каждой пары рядов распределения вычисляют математическое ожидание (М) числа инверсий (u):
где m и n - число наблюдений в каждом ряду, и среднее квадратическое
Приняв уровень значимости критерия q=5%, рассчитывают область больших по абсолютной величине отклонений:
и определяют положение полученного значения числа инверсий относительно этой области.
Важное преимущество непараметрического критерия Вилкоксона состоит в том, что он применим при обработке экспериментальных данных как соответствующих нормальному распределению, так и отклоняющихся от него.
АДАПТАЦИЯ РЫБ К ЯДАМ. КУМУЛЯЦИОННЫЙ ЭФФЕКТ
Изучение адаптации рыб к ядам промышленных сточных вод представляет исключительный теоретический и практический интерес в связи с хроническим действием малых концентраций токсических веществ в большинстве водоемов.
Адаптацией называют приспособление организма к какому-либо стрессирующему фактору внешней среды. На первый план здесь выступает необходимость сохранения физиологической нормы, лежащей в основе биологической нормы - сохранения вида в целом.
Глубина повреждения и выраженность сдвигов со стороны различных функциональных систем организма под влиянием ядов промышленных сточных вод зависит от концентрации и природы яда. Патологические изменения, не затронувшие основные функции, могут иметь временный характер и не обязательно приведут рыбу к гибели, хотя при этом она может лишиться хозяйственной ценности (например, порча вкуса или запаха, угнетение роста и т. д.). Отсюда понятна необходимость исследования потенциальных возможностей адаптации рыб к ядам для прогноза результатов длительного контакта рыб с субтоксическими концентрациями ядов в водоеме.
У насекомых и клещей, например, довольно быстро возникает адаптация к органическим ядам, что затрудняет борьбу с вредными видами этих организмов.
Вработах Н. С. Строганова и Л. В. Колосовой (1962), а также Н. С. Строганова (1964) представлены результаты опытов, направленные на выявление возможности адаптации Daphnia magna к гексахлорбутадиенту (ГХБ). На первом этапе контакта дафний с ядом численность потомства в растворах ГХБ 0,015-0,5 мг/л уменьшалась, но численность последующих 3-4 генераций дафний достигла контрольного уровня. Механизм этого феномена остается пока неясным, идет ли речь об отборе устойчивых к ГХБ (равно как и к любому другому яду) особей или имеет место истинная адаптация дафний к яду.
Вопытах Н. С. Строганова, проведенных совместно с Т. Е. Шошиной, установлено, что адаптация дафний
кцинку и кадмию так и не наступила в течение трех генераций, на протяжении которых , снижение численности особей нарастало.
Адаптацию рыб к ядам изучали редко, имеются буквально единичные сообщения по этому поводу. В опытах Д. Гудмана (Goodman, 1951) и Р. Аффлека (Affleck, 1952) с цинком, а Р. Паули (Paul, 1952) с медью было отмечено, что предварительный контакт рыб с этими ядами может оказать влияние на время выживания рыб в токсических растворах, заметно повысив его.
По свидетельству М. М. Телитченко, рыбы, не погибшие в растворах той или иной концентрации стронция, урана или тория, могут длительное время находиться в растворах этих веществ при концентрациях в десятки раз превышающих исходные.
Обнаружено, что устойчивость рыб к некоторым дезинфикато-рам увеличивается по мере нахождения в
50